[0001] Title: A process for the gene manipulation of plant cells, recombinant plasmids, recombinant bacteria, plants
[0002]This invention relates to a process for the gene manipulation of plant cells, which comprises introducing foreign DNA into plant cells by infecting the plant cells with one or more recombinant Agrobacterium tumefaciens strains, which contain foreign DNA to be transferred to the plant cells between the left- and right-hand ends of T-DNA necessary for such a transfer and are capable of transferring this foreign DNA to the. lant cells.
[0003]The invention further relates to a recombinant plasmid consisting of a bacterial vector plasmid and at least one DNA insert, and to a recombinant Agrobacterium tumefaciens strain, which contains foreign DNA to be transferred to plant cells between the left- and right-hand ends of T-DNA necessary for such a transfer and is capable of transferring this foreign DNA to the plant cells.
[0004]Finally, the invention also relates to the plants deriving from the transformed cells and to the parts consisting of one or more cells or products thereof.
[0005]The AgrPfracteriuro t mefaciens technology for the gene manipulation of plants, which technology has strongly developed and has been applied on a large scale since its first discovery, is in principle still limited, despite all progress made through the years, to the application in specific, substantially dicotyl plant species, such as tobacco, tomato, colza etc. Up to the present time a successful transformation of monocotyl plant species has essentially only been described for some Liliaceae, namely Asparagus (Bytebier et al, Proc. Natl. Acad. Sci. USA £4., 5345-5349, 1987) and Narcissus (Hooykaas et al, Nature 211., 763-764, 1984), some Iridaceae (Graves, et al, J. Bacteriol. 169. 1745-1746, 1987), some Dioscoreaceae (Schafer et al. Nature 327. 529-532, 1987), and some Gramineae, namely Zea mays (Graves and Goldman, Plant Mol. Biol. 1, 43-50, 1986) and Oryza sativa (Raineri et al, Biotechnology ϋ, 33-38, 1990) .
Apart from these exceptions, one has not yet succeeded in realizing by means of the Agrobacterium tumefaciens technology an efficient transformation of onocotyi plant species, including the important cereals such as maize, wheat, barley, rye, oat, rice etc. and in producing stably transformed monocotyi plants. In a broader sense this applies to many species of plants, not only to the cereals belonging to the group of monocotyi plants but also to grasses such as ryegrass and fescue, leguminous plants such as leek, onion and garlic, ornamental plants such as tulipe, hyacinth, bromeliad, iris, orchid etc., and other plant species such as conifer etc.
[0006]There seem to be indications that the problem is not due to the transfer of the DNA. Thus it has been found possible to transmit viral genomes encoded in the form of DNA by means of Agrobacterium tumefaciens to maize (Grimsley et al.
[0007]Biotechnology £, 185-189, 1988) and wheat (Dale et al, Seventh International Wheat Genetics Symposium 1, 719-722, 1988) . Perhaps the problem is therefore due to the integration of the DNA in the genome: the integration system which Agrobacterium tumefaciens bacteria successfully use in dicotyl plants might not or not efficiently work in monocotyi plants. Generally, however, it is assumed that a poor transfer of the T-DNA to the plant is responsible for the failure of the technology in monocotyi plants . In order yet to obtain the desired transformation in monocotyi plants, different alternative transformation techniques have been conceived and developed in which no use is made of Agrobacterium tumefaciens bacteria for transmitting DNA to the plant cell. Examples thereof are microinjection with DNA, bombardment with DNA-carrying particles, electroporation of protoplasts with DNA etc.
[0008]These techniques have been successfully employed for the introduction of DNA into plant cells, but generally only a passing expression of the genes introduced could be observed. As regards the use of integration systems, the selection made so far concerns either homologous genetic recombination.
which is a hardly efficient system and remains limited in practice to cells in meiosis, or transposition, which is an efficient system and is also active during the normal cell cycle but has the drawback that the genes introduced are not stable in the first generation and that it is necessary to wait one or more generations before the transposon and the genes introduced via transposition are separated by mendelizing out.
[0009]Transposons are, in fact, genetic elements which can move within the genome of the host cells in which they are normally present. They have virtual ends, repeated border sequences which are in general invertedly repeated and may be rather different in length. At the location of their insertion into the genome a piece of host sequence is found back directly repeated to the left and the right of the transposon sequence. There is generally no specific selection of the location of insertion. In case of excision either the original sequence is restored or a deletion occurs or part of the sequence remains at the relevant location (insertion) . In its simplest form a transposon consists of a transposase gene located between terminal sequences which contain transposase binding sites and optional binding sites for other host enzymes and terminate with the repeated border sequence. These terminal sequences, together with the directly repeated sequences of the target DNA to the left- and right-hand of the border sequence, will also be referred to hereinbelow as left- and right-hand transposon ends .
[0010]The present invention provides a process which removes the main objections of the known methods and gives a very efficient and stable transformation of cells of monocotyi plants, which has not been found possible so far. For this purpose use is made according to the invention of the efficient transfer system of Agrobacterium tumefaciens bacteria in combination with the efficient integration system of transposons.
The invention relates in a first aspect to a process for the gene manipulation of cells of plants which, in essence, cannot be transformed by Agrobacterium tumefaciens with integration by T-DNA, with foreign DNA being introduced into the plant cells by infecting the plant cells with one or more recombinant Agrobacterium tumefaciens strains which contain foreign DNA to be transferred to the plant cells between the left- and right-hand ends of T-DNA necessary for such a transfer and are capable of transferring this foreign DNA to the plant cells, the foreign DNA located between the left- and right-hand ends of T-DNA comprising:
[0011](A) a transposase gene located in an expression cassette active in the plant cells but not between the left- and right- hand transposon ends necessary for integration in the DNA of the plant cells, and
[0012](B) a recombinant transposon comprising the left- and right- hand transposon ends necessary for integration in the DNA of the plant cells with an intermediate DNA fragment to be integrated in the DNA of the plant cells but containing no DNA fragment which can lead in the plant cells to expression of an active transposase.
[0013]By the words "plants that, in essence, cannot be transformed by Agrobacterium tumefaciens with integration by
[0014]T-DNA" are meant all those plant species in which Agrobacterium tumefaciens is not capable of effecting an efficient integration of the T-DNA in the plant genome, which roughly means almost all the non-dicotyl plants . The invention is therefore eminently suited for the gene manipulatin of monocotyi plants and other non-dicotyl plants, preference being given according to the invention to gene manipulation of plant cells of cereals such as maize, wheat, barley, rye, oat, rice etc., grasses such as ryegrass and fescue, leguminous plants such as leek, onion and garlic, ornamental plants such as tulipe, hyacinth etc. and other plant species such as conifer etc.
According to the invention there is used a transposase gene which is not located between the left- and right-hand transposon ends necessary for integration in the DNA of the plant cells. According to the invention there is further used a recombinant transposon containing no DNA fragment which can lead in the plant cells to expression of an active transposase. This means that the recombinant transposon is an inactive transposon. Consequently, a desirable high stability of the transformed plants can be obtained. The drawback of instability -normally connected with the use of transposons is, in fact, the result of the presence of a complete (or active) transposon, i.e. a transposon with a transposase gene located between the transposon ends . This gene is then co-incorporated with the transposon in the plant genome and may therefore give rise for several generations to the undesirable instability. According to the present invention there is used an active transposase gene, namely in order to ensure efficient integration of the foreign DNA located in a transposon in the DNA of the plant cell (in the plant cell the transposase gene leads to the formation of transposase which, in cooperation with nuclear factors of the plan , is capable of effecting efficient integration of the transposon in the genome) but if this transposase gene is not within a transposon, in conformity with the preferred embodiment, it is not itself efficiently integrated in the genome in case of transformation of non-dicotylidons, such as in particular monocotylidons . This means that the desired stability is obtained already in the first generation of plants.
[0015]The transposase gene preferably consists of a genomic clone of the transposase gene, i.e. of genomic DNA which comprises not only the encoding exons but also the introns normally present . Although the use of a cDNA coding for the transposase is also possible, practice has shown that it gives a substantially less efficient integration of the recombinant transposon. The use of a genomic transposase gene does not mean that the natural promoter of the transposase gene should
be used too. Practice has shown that it is very advantageous to use, instead of the natural promoter, another promoter active in plants, in particular a promoter that can effect efficient expression in cells from which germinative cells develop. A promoter suitable for this purpose is the known CaMV 35S promoter. The use of the pTRl* or pTR2 ' promoter is also posible and favorable if, as is also preferably done according to the invention, transfer occurs by microinjection with the apical meristem being mechanically damaged. As a result, the wound-inducible regulators of gene expression for these promoters are present in the meristematic cells. The use of such a construct of a genomic transposase gene under control of another promoter such as the CaMV 35S, the pTRl' or the pTR2 ' promoter, therefore constitutes a highly preferred embodiment of the invention.
[0016]According to the invention it is particularly preferred that between the left- and right-hand transposon ends the recombinant transposon comprises at least one foreign gene located in an expression cassette active in the plant cells. By this is meant any gene that is to be incorporated in the genome of the plant for whatever reason. It may be a gene coding for a desirable property of the plant, such as resistance to herbicides, to pathogens, such as specific fungi and insects, or to environmental factors, or properties, such as (in ornamental plant cultivation) a specific flower color, flower scent, flower size etc., or properties such as (in case of food crop) better nutritional properties, a higher yield etc. It may also be a gene coding for a substance one wishes to produce on a large scale by cultivating a plant capable of forming the substance by means of gene manipulation. The gene to be introduced into the plant may therefore be a gene within the scope of plant breeding or a gene within the scope of biological production of specific products (proteins and enzymatic products, DNA, RNA etc.) . It is also possible that between the left- and right- hand transposon ends the recombinant transposon contains a
promoter active in plants, instead of a foreign gene in an expression cassette active in plants. Consequently, it becomes possible to alter the expression behavior of specific genes and thus the phenotype. It is further preferred that also one or more selectable marker genes suitable for selection of transformed bacteria and located in an expression cassette active in the bacteria are located between the left- and right-hand ends of T-DNA. This enables selection of Agrobacterium tumefaciens bacteria with the desired DNA constructs . Marker genes suitable for this purpose are known to those skilled in the art. These are often genes the expression of which leads to a resistance to specific antibiotics, such as resistance to kanamycin, chloramphenicol, ampicillin, tetracycline, spectinomycin etc. It is particularly preferred here that one or more selectable marker genes suitable for selection of transformed bacteria and located in an expression cassette active in bacteria form part of the recombinant transposon. This preferred embodiment has the advantage that the marker genes coupled to the foreign gene are co-incorporated in the genome of the plant so that in cases that as a result of the incorporation of the recombinant transposon in a gene of the host the transformed plant shows another phenotype (which is not due to the introduced genetic information as such) or that the foreign gene introduced proves to function differently in later generations than in the first generations (e.g. as a result of mutation) the recombinant transposon can be cloned for further examination from the plant genome into bacteria again by means of selection with the marker gene active in bacteria.
[0017]It is further preferred that also one or more selectable marker genes suitable for selection of transformed plant cells and located in an expression cassette active in the plant cells are located between the left- and right-hand ends of T-DNA. Also here it applies that many marker genes suitable for this purpose are known to those skilled in the art . Thus
the resistance to, e.g., antibiotics or herbicides can be mentioned as a property which, as is known to those skilled in the art, can be used for a selection of successfully transformed plants . It is particularly preferred here that one or more selectable marker genes suitable for selection of transformed plant cells and located in an expression cassette active in the plant cells form part of the recombinant transposon. This preferred embodiment has the advantage that the marker genes are co-incorporated in the genome of the plant so that also later generations can be selected on the basis of the properties encoded by the marker genes.
[0018]A first variant of the process according to the invention is characterized in that the plant cells are infected with a mixture of two different recombinant A.. tumefaciens strains, one of which comprises between the left- and right-hand ends of T-DNA the transposase gene located in an expression cassette active in the plant cells and the other the recombinant transposon between the left- and right-hand ends of T-DNA. This variant has the advantage of having a wide range of applications, i.e. there can always be used the same Agrobacterium strain supplying the transposase gene and only the second strain containing the recombinant transposon must be constructed for each new gene. In this second strain there is used a construct in which a multiple cloning site is located between the transposon ends so as to have a great freedom of choice with respect to the DNA fragments to be incorporated in the transposon.
[0019]In practice, in this first variant according to the invention both the first and the second A. tumefaciens strain will comprise between the left- and right-hand ends of T-DNA at least one marker gene for selection of transformed bacteria and at least one marker gene for selection of transformed plant cells. It is preferred here that at least the marker genes for selection of transformed plant cells which carry the two bacterial strains within the T-DNA ends are different from
each other so as to recognize plant cells successfully transformed by the two strains.
[0020]An alternative embodiment of the process according to the invention is characterized in that the plant cells are infected with one recombinant A. tumefaciens strain which comprises between the left- and right-hand ends of T-DNA both the transposase gene located in an expression cassette active in the plant cells and the recombinant transposon. This variant has the advantage of more chance of success of the transformation, i.e. the desired DNA transfer and integration will have taken place in a larger part of the treated plant cells .
[0021]In practice, in this second variant according to the invention the A., tumefaciens strain will comprise between the left- and right-hand ends of T-DNA at least one marker gene for selection of transformed bacteria and at least one marker gene for selection of transformed plant cells.
[0022]The process according to the invention is of special importance to the transformation of plant cells of a monocotyi plant species such as maize, wheat, barley, rye, oat, rice etc. An efficient transformation of such crops has not been found possible so far.
[0023]The invention can also be used, however, for the purpose of altering phenotypic characteristics of plants resulting from the presence of inactive transposons in one or more genes of the plant by removing such a transposon from the gene in which it is contained. For this purpose the invention provides a process for the gene manipulation of cells of plants which, in essence, cannot be transformed by Agrobacterium tumefaciens with integration by T-DNA, which comprises introducing foreign DNA into the plant cells by infecting the plant cells with a recombinant Agrobacterium tumefaciens strain which contains foreign DNA to be transferred to the plant cells between the left- and right-hand ends of T-DNA necessary for such a transfer and is capable of transferring this foreign DNA to the plant cells, which process is characterized in that plant

[0024]hand transposon ends necessary for integration in the DNA of plant cells.
[0025]The invention further relates to a recombinant plasmid consisting of a bacterial vector plasmid and at least one DNA insert, which recombinant plasmid is characterized in that at least one DNA insert comprises a recombinant transposon which contains the left- and right-hand transposon ends necessary for integration in the DNA of plant cells but no DNA fragment which can lead in plant cells to expression of an active transposase. .
[0026]A preferred embodiment of such a recombinant plasmid is characterized in that between the left- and right-hand transposon ends the recombinant transposon comprises one or more restriction endonuclease recognition sequences suitable as cloning site. Such a plasmid has a wide range of applications as a carrier for genes that are to be introduced into the genome of plants.
[0027]A special preferred embodiment of such plasmids is characterized in that the recombinant transposon comprises between the left- and right-hand transposon ends at least one foreign gene located in an expression cassette active in plant cells.
[0028]In such recombinant plasmids it is further preferred that the recombinant transposon comprises between the left- and right-hand transposon ends one or more selectable marker genes suitable for selection of transformed plant cells and located in an expression cassette active in plant cells, and that the recombinant transposon comprises between the left- and right-hand transposon ends one or more selectable marker genes suitable for selection of transformed bacteria and located in an expression cassette active in bacteria.
[0029]In yet another aspect the invention provides a recombinant Agrobacterium tumefaciens strain which contains foreign DNA to be transferred to plant cells between the left- and right-hand ends of T-DNA necessary for such a transfer and is capable of transferring this foreign DNA to the plant
t
[0030]
[0031]The invention is finally also contained in plants and parts consisting of one or more cells or products thereof, which plants derive from plant cells transformed by the process according to the invention. By parts and products of plants is meant, e.g., the bulb, the tuber or the root, the flowers, the seed and the meal made therefrom etc.
[0032]The invention will be further explained with reference to the accompanying drawings, which diagrammetically show in Fig. 1 a T-DNA construct comprising both a foreign gene X included in an inactive transposon and a transposase gene not located in an active transposon.
[0033]Fig. 2a a T-DNA construct comprising a foreign gene X included in an inactive transposon.
[0034]Fig. 2b a T-DNA construct which can be used in combination with that of Fig. 2a and comprises a transposase gene not located in an active transposon.
[0035]Fig. 3a a T-DNA construct comprising a foreign gene X included in an inactive transposon which comprises a marker gene interrupted by a second inactive transposon for selecting transformed plant cells.
[0036]Fig. 3b a T-DNA construct which can be used in combination with that of Fig. 3a and comprises a transposase gene not located in an active transposon,
[0037]Fig. 4a a T-DNA construct comprising a foreign gene X included in an inactive transposon, which foreign gene X is interrupted by a second inactive transposon,
[0038]Fig. 4b a T-DNA construct which can be used in combination with that of Fig. 4a and comprises a transposase gene not located in an active transposon, Fig. 5a the structure of the recombinant plasmid pMHIO, pMHIO-LTS and pMHIO-DsB, respectively a plasmid having a complete transposon, a transposase gene with only the left transposon end and a transposon without transposase gene, Fig. 5b an example of a T-DNA construct with plasmid pMHIO-LTS according to the invention,
Fig. 6a the structure of the recombinant plasmid pMH10Dsl5GUS,
[0039]Fig. 6b an example of a T-DNA construct with this plasmid according to the invention, Fig. 7 the structure of the recombinant plasmid pMH10Ds34CAT,
[0040]Fig. 8 the structure of the recombinant plasmid pMH10Dsl2lAA,
[0041]Fig. 9 the structure of the recombinant plasmid pMHHDs4IPTkan,
[0042]Fig. 10 the structure of the recombinant plasmid pϋCSSAc9Dsl9, suitable for cloning foreign genes between the transposon ends of Ac9,
[0043]Fig. 11a the structure of the recombinant plasmid pUCSS19tpnAc9P35S,
[0044]Fig. lib the structure of the recombinant plasmid pUCSS19tpnAc9PTR1 ,
[0045]Fig. lie a T-DNA construct with the recombinant plasmid pϋCSS19tpnAc9P35S according to the invention. Fig. 12a the structure of the recombinant plasmid pϋCSSDsl9BTKA,
[0046]Fig. 12b a T-DNA construct with this plasmid according to the invention,
[0047]Fig. 13a a cointegrate plasmid with the constructs pMHHDs IPTkan and pUCtpnAc9P35S, and
[0048]Fig. 13b an example of a T-DNA construct with the cointegrate plasmid pMHHDs4IPTkan/pUCtpnAc9P35S.
[0049]In the drawings the symbols have the following meanings: X represents a foreign gene located in an expression cassette, not separately shown, active in plant cells; if the gene is interrupted by an insert, the component parts of the gene are indicated by X' and X", respectively;
[0050]SI represents a marker gene which can be used for the selection of transformed plant cells; here, too, the component parts are indicated by SI' and SI", respectively, when the gene is interrupted by an insert;
S2 and S3 also represent marker genes suitable for the selection of successfully transformed plant cells, further referred to as plant selection genes; si and s2 represent marker genes suitable for the selection of transformed bacteria, further referred to as bacteria selection genes; tpn represents a genomic clone of a transposase gene located in an expression cassette, not separately shown, which can function in plant cells; LTA and- RTA represent respectively the left end of T-DNA and the right end of T-DNA, which ends are necessary for transferring the T-DNA by the Agrobacterium tumefaciens bacteria to the plant cell;
[0051]LTS and RTS represent respectively the left end of a transposon and the right end of an active or inactive transposon, ; x represents the sequence of the waxy gene of maize from which Ac9 was cloned;
[0052]Fig. 1 shows a T-DNA construct according to the invention, one plasmid containing between the T-DNA ends LTA and RTA both an inactive transposon having a foreign gene X and a plant selection gene SI contained therein and a transposase gene tpn not located on an active transposon. The selection genes shown in Fig. 1 are facultative, for that matter, in particular one of the bacteria selection genes and one of the plant selection genes being dispensable, if required. As regards the transposase gene, it is important that it is not located within the transposon ends LTS and RTS. This does not mean that the transposon ends must be completely absent: often removal of (a significant portion of) one of the transposon ends will be sufficient to prevent efficient incorporation of the transposase gene in the genome of the transformed plant cells. Thus, for instance, one transposon end could be retained in Fig. 1 beside the transposase gene (see Fig. 5a) .

[0053]
[0055]
[0056]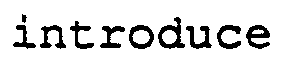 s (Ds) derive
s (Ds) derive

[0057]1 into the genome vity of this ations because of a gene) or to Ac insertion
[0058]1 ibed here the Ac insertion ize. In conformity erium is used o cells of the
[0059]2 he PstI clone of hlman et al, 1984, agment of plasmid

 al ori sequence
al ori sequence
[0060]25 for replication in gram-negative bacteria, but a non¬ functional amp and tet resistance gene and moreover a BamHI-
[0061]strain containing the Ti plasmid pGV3850 (Zambryski et al, 1983), in which pBR322 is cloned as a Hindlll fragment into the T-DNA of pTi CSS from which, furthermore, all Hindlll fragments were cut, except the Left Border (LB) fragment (Hindi11-10) and the Right Border (RB) fragment (HindIII-23) containing the nopaline synthase gene (ηos) . The plasmid pGV3850 was obtained from Laboratorium voor Genetika RUG, Ledeganckstraat 35, B9000 Ghent, Belgium.
[0062]The method of conjugation was described by Van Haute et al (1983) . By. selection of transconjugants for rif. str/spc and amp resistance a cointegrate was selected, pMHIOlLTS being integrated between the borders of the T-DNA.
[0063]Maize lines obtained by inbreeding for at least ten years were made available, in the form of seed, by the firm of Clovis Matton, Avelgem-Kerkhove, Belgium. These lines contain no active transposon Ac, which, inter alia, could be proved by breaking down the DNA of young leaves with PvuII and hybridizing it with the internal Hindlll fragment of Ac (Ac9(wx) : bp: 1270-2877) : no hybridizing band was observed corresponding to the 2626 bp internal PvuII fragment of Ac
[0064](Ac9(wx) : bp: 719-3345); different bands were found indeed but the smallest band exceeded 4 kb. Crossings with test lines for Ac activity have never shown transposition phenomena in the lines used. The lines are used for the production of commercial hybrids .
[0065]Seeds of ten maize lines were disinfected with NaCIO 2% and then laid on sterile wet sand to germinate on sterile sand at 25°C in the dark. Three days after germination 50 nl induced £. turn, culture were integrated with the construction pMHIO LTS by genetic recombinantion between the borders of the T-DNA (Fig. 5b), injected into the apical meristem by microinjection.
[0066]The induction medium has been described by Vernade et al (1988) . After one week at 20°C the young plants were cultured together with untreated plants in large pots in an air- conditioned greenhouse in a mixture of compost and sandy clay
in their normal growing season (May to October) . Each plant was fertilized manually, both the plume and the ear being kept in small bags. Each plant was individually watered in the pot. The ears were harvested in September and October and after drying the seed was stored in paper bags during winter at 6°C. The next year the plants were sown on ear lines in the soil in the greenhouse and again fertilized manually and evaluated by corn growers for their characteristics both during growth and in surmaturity. The following improvements over the original lines were observed in some ear lines :
[0067]- plume more branched
[0068]- 30 - 50 cm higher plants
[0069]- stronger stem - significant increase in the number of grain rows per ear
[0070]- remains green for a longer time
[0071]- absence of fusariosis
[0072]- better anchoring by secondary roots
[0073]Seeds of plants having deviating features as well as of control plants were germinated in the laboratory under sterile conditions and DNA was prepared from the young leaves, after which the plants were planted in the greenhouse in large pots,
[0074]The DNA of each plant was then broken down such as for RFLP mapping with different restriction enzymes and hybridized with the internal Hindlll fragment of Ac9. The control plants showed the same band pattern, as regards the prominent bands, as the original line. In plants with deviating features either disappearance of one or more fragments hybridizing with the DNA probe or the disappearance of a fragment and the appearance of a new fragment with a larger molecular weight was observed.
[0075]Example 2 (comparative example)
[0076]Insertion of a foreign gene into the genome of maize by using Agrobacterium tumefaciens as transfer system and transposon Ac as integration system.
For the purpose of comparison an active transposon Ac9(wx) is used here. By using the integration properties of the transposon Ac and the transfer properties of A., turn, a foreign gene located between the recognition sequences of the transposon can be incorporated in the genome of maize by means of transposase of the same transposon.
[0077]As described, two A. tumefaciens strains are used by way of example. The first contains the β-glucuronidase gene of Escherichia coli located between the left- and right-hand transposon ends of Ac9 (wx) , which, in turn, is located between the Left and Right Border (LB and RB) of the T-DNA on a non- oncogenic Ti plasmid. The second contains a complete transposon Ac9(wx) located between LB and RB of the T-DNA. The A. turn, strain containing the transposon Ac9 in the T-DNA is shown in Fig. 5a. The other A,, turn, strain was obtained in the following manner:
[0078]Plasmid pBI 221 contains the β-glucuronidase gene of £. coli (ϋidAJ under the control of the CaMV 35S promoter (Cauliflower Mosaic Virus 35S RNA promoter) and the mRNA distal end of the nopaline synthase gene of the Ti plasmid of A. turn. T37. This plasmid was available from Dr R. Jefferson, c/o AFRC Institute, Colney Lane, Norwich NR4 7UA, U.K.
[0079]The plasmid pMHIO shown in Fig. 5a was cut with EcoRI and partially with flindlll and the 10.1 kb fragment was ligated with the 3 kb EcoRI-Hindlll fragment of pBI 221 containing the sequence of the chimeric uidA gene of this last plasmid. The resulting plasmid pMH10Dsl5GUS contains the chimeric uidA gene in a transcription orientation contrary to that of the (partially) deleted transposase gene.
[0080]&- turn, bacteria with this plasmid, introduced in the manner described in Example 1, show β-glucuronidase activity. The construction is shown in Figs. 6a and 6 .
[0081]The same maize lines described in Example 1 were used for the transformation. In three-day-old germs the apical meristem was infected by microinjection with 50 nl of a
mixture of the two above-described ., turn, strains induced for transfer. In a first phase some hundreds of germs were tested after 1 or 2 weeks for β-glucuronidase activity by incubating a piece of tissue containing the apical meristem with X-gluc (5-bromo-4-chloro-3-indolyl-D-glucuronide) . In about 7.5% of the tissues blue staining occurred. ith a dark field microscope it was clearly visible that the cells in the tissue cuts and not the remaining bacteria were the origin of the purple color. In particular small cells around the vascular bundles were stained. With an electron microscope the precipitate of dimeric bromo-chloro indole was found in cells which were removed from the place of injection up to ten cell layers . Sporadically, cut bacteria were visible which were only present in the extracellular spaces but showed no precipitate.
[0082]Other treated germs were grown in the greenhouse in a normal growing season, manually fertilized and the seed was harvested as described in Example 1.
[0083]During the winter months these seeds were germinated in sterile conditions and the proteins were isolated from the young leaves and subjected to polyacrylamide gel electrophoresis (PAGE) under non-denaturing conditions. Beta- glucuronidase activity was detected by covering the gel with a thin agarose gel containing the substrate methyl-umbelliferyl- D-glucuronide which fluoresces in UV light if the β- glucuronidase enzyme causes release of methyl umbelliferone. A fluorescent band was found in about 1% of the tested plants . DNA was prepared from leaves and pieces of roots of these plants and, as in RFLP mapping, hybridized with a 2.9 kb Hindlll-EcoRI fragment containing the chimeric uidA gene. In the lane of the agarose gel containing maize DNA broken down with Hindlll and EcoRI hybridization with the probe was found on a level with the place where about 3 kb fragments migrate. Of young Fl plants pieces of leaves and roots were incubated in X-gluc as described by Jefferson (GUS gene fusion system user's manual, 1987) . Blue stains, zones and sectors
were found in the pieces of tissues of PAGE-β-glucuronidase positive plants: around the vascular bundles, around the stomata, V-shaped segments of the roots, etc.
[0084]Light microscopic and electron microscopic analyses confirmed the expression of the β-glucuronidase enzyme in the plant cells.
[0085]This example clearly shows that by using an active transposon in combination with a chimeric transposon containing a foreign gene but no transposase gene between the transposon ends expression is only obtained in specific cells still containing the chimeric transposon, while most of the plant cells show no expression. The same phenomenon can be observed after transformation of plants such as tobacco which are efficiently transformed by Agrobacterium but in this case instability occurs after transformation with a chimeric transposon, both in the presence of an active transposon and in the presence of a transposase gene. This shows the difference between plants in which Agrobacterium can efficiently integrate foreign DNA in the plant genome and plants in which Agrobacterium cannot.
[0087]Generation of mutations in barley by using a transposase gene and a chimeric transposon.
[0088]In this example the mutagenic properties of a transposon are used by integrating a chimeric transposon containing a plant selection gene but no transposase gene in the genome of barley by means of a transposase gene that is not located between transposon ends.
[0089]Use is made of two Agrobacterium strains for the transfer: the first strain contains the chimeric transposon between the T-DNA ends and the second strain contains the transposase gene between the T-DNA ends. This example corresponds with Figs. 3a and 3b.
The employed plasmids are pMH10Dsl5GUS and pMHIOLTS. As described before, they are conjugated from Escherichia coli to Agrobacterium and integration in pGV3850 selected by also adding spectinomycin to the medium. Seeds of two- and six-rowed winter barley ear lines (F7-F8) were supplied by the plant breeding station of the firm of Clovis Matton, Avelgem-Kerkhove, Belgium.
[0090]Seeds of 12 ear lines were disinfected with 6% NaCIO and the seeds laid on moist sand to germinate at 4°C. Germinating seeds were further incubated at room temperature under sterile conditions .
[0091]In 3-5-day-old seedlings the apical meristem was infected by microinjection with Agrobacterium strains containing pMH10Dsl5GUS or pMHIOLTS. Transfer was induced as described by Vernade et al (1988) . Infections were conducted both with the strain containing pMG15 (control) and with the strain containing pMHIOLTS (control) or with both of them (experiment) . About 300 seedlings were treated per ear line: a hundred per experiment. Further control experiments were conducted on tobacco, for rapid tests of expression.
[0092]After 1 week at 22°C the young plants were cultured further in a greenhouse and the seed was harvested after self- pollination.
[0093]In the next generation the plants were sown again in a greenhouse and selected by barley growers for deviating features .
[0094]The following variants were traced:
[0095]- difference in height of the plant during growth
[0096]- difference in time of growth - mutations from erectant to nutant or conversely
[0097]- difference in number of grains per ear
[0098]- difference in flattening
[0099]- better or poorer phytosanitary condition.
[0100]Of 37 ear lines with clear morphological or phytosanitary differences and 110 other ear lines with less
clear deviations five ripe ears were further examined for expression of the uidA gene in the chimeric transposon.
[0101]Nearly all the 37 plants with clear differences over the original ear line express the uidA gene to a greater or lesser degree. No β-glucuronidase activity could be demonstrated in the controls, except in the case of bacterial infection. Only 9 plants with less clear deviations show uidA expression.
[0102]In tobacco β-glucuronidase activity was found when the gene was present in the infection, with or without additional infection with the transposase gene.
[0104]Construction of recombinant plasmids with plant selection genes or genes with industrial applications between left- and right-hand transposon ends in the plasmid pMHIO.
[0105]In this example the construction of some plant selection genes was shown by using unique restriction sites in the sequence of Ac9 in the plasmid pMHIO. The chloramphenicol acetyl transferase gene in an expression cassette consisting of the 35S promoter of CaMV and the no_s_-terminator of the T-DNA of Agrobacterium tumefaciens is available as plasmid pCAMVCN from Pharmacia-LKB, Bromma, Sweden. The complete chimeric gene was cut as Xbal fragment. The plasmid pMHIO was partially cut with Bell (position 1530 in Ac9Wx) and BamHI (position 4498 in Ac9Wx) and the 8.1 kb fragment was ligated back. This plasmid, called pMHIODsB, contains unique restriction sites in the remaining Ac9 sequence: Nsil (five sites at position 571, 972, 1003, 1183, and 1251 of Ac9Wx) , Xhol (at position 1125 of Ac9Wx) and Xbal (at position 1257 of Ac9Wx) (Fig. 5a) . The cat expression cassette was cloned into the unique Xbal site of Ac9. The resulting plasmid was called pMH10Ds34CAT (Fig. 7), conjugated to Agrobacterium and integrated in the plasmid pGV3850. By transformation of tobacco the expression of the cat gene was proved.
The genes iaaM and iaaH from the T-DNA of Agrobacterium tumefaciens Ach5 (Gielen et al, 1984) were cloned from the plasmids pGV814 (EcoRl-Clal clone of iaaM) (Budar et al, 1986) and pGV824 (Hindlll clone of iaaH) (Budar et al, 1986) . The complete construct with the genes iaaM and iaaH was first cloned into pBR322 (Bolivar et al, 1977) by a threefold ligation of the 3.6 kb pBR322 Pstl-Clal fragment, the 3.7 kb EcoRl-Clal fragment of pGV814 and the 2.7 kb partial Pstl-EcoRl fragment of pGV824. From this was cloned the 6.4 kb PstI fragment in the Nsil sites (position 571 and 2881 of
[0106]Ac9Wx) of pMHIO. The resulting plasmid pMH10Dsl2IAA (Fig. 8) was conjugated to Agrobacterium and integrated in the plasmid pGV3850 by additional selection with spectinomycin. The usability of the construction was tested by transformation of tobacco and expression of the iaa genes.
[0107]The gene ipt from the T-DNA of Agrobacterium tumefaciens C58 together with the bacterial nptll gene of transposon Tn5 of plasmid pG4L (Inze et al, 1984) was cloned as a BamHI fragment in pUC19 and isolated therefrom as a partial Sphl-EcoRl fragment of 3.7 kb and cloned into pMHll cut with EcoRl and partially with Sphl. pMHll is a derivative of pMHIO obtained by cutting in the vector plasmid of pMHIO the 0.8 kb Sphl fragment located between the str/spc resistance gene and the (partially deleted) tet resistance gene, blunt ending the ends with SI nuclease and ligating back the vector. The PstI clone of Ac9Wx was again incorporated in the resulting vector in the unique PstI site. The orientation, however, is different from that in pMHIO.
[0108]The resulting plasmid pMHHDs4IPTkan (Fig. 9) was conjugated to Agrobacterium and integrated in pGV3850. by additional selection with spectinomycin and then with kanamycin. The usability of the construction was proved by transformation of tobacco and expression of the gene ipt .
[0109]Example 5
Construction of a recombinant transposon with a multiple cloning site between the transposon ends .
[0110]The Sphl-Kpnl fragment from mini-Sa, a derivative of pR702 (Leemans et al, 1983) containing the sm-sp adenosyl transferase gene encoding resistance to streptomycin and spectinomycin was blunt ended with T4 DNA polymerase and cloned into the plasmid pUC19 (Yanisch-Perron et al, 1985) , blunt end cut with Sspl. This plasmid was 4330bp and will be referred to hereinbelow as pUCSS19.
[0111]Both orientations of the sm/sp resistance gene are suitable. However, the work was further done with the plasmid in which the amp resistance gene and the sm/sp resistance gene have the same transcription orientation. pUCSS19 was blunt end cut with PvuII and both the 4030bρ vector fragment and the 300bp MCS fragment were separated and stored.
[0112]The PstI clone of Wx-Ac9 was cut from pMHIO (Fig. 5a) and blunt ended with T4 DNA polymerase. This fragment was cloned into the 4030bp PvuII fragment of pUSS19. This plasmid was 8840bp and will be referred to hereinbelow as pUCSSAc9. pUCSSAc9 was cut with Nsil and MscI and the Nsil end was blunt ended with T4 DNA polymerase. This fragment was 4980bp and contained the ends of transposon Ac9Wx. Into this was cloned the 300bp PvuII fragment of pUC19. One of the orientations is shown in Fig. 10. The plasmid was 5280 bp and will be referred to as pUCSSAcDsl9. It can be used for cloning numerous genes with their expression cassette. It can be conjugated to Agrobacterium with plasmid pGV3850 as described in the first example by additional selection with spectinomycin.
[0114]Construction of recombinant plasmids containing plant selection genes or genes with industrial applications and located between the transposon ends of pUCSSAcDsl9.
The delta endotoxin gene of the 42 MDa plasmid of Bacillus thyriπ jeπsis var. Berliner 1715 (Klier et al, 1982), the sequence of which was published by Hδfte et al, 1986, was obtained from S.A. Solvay, Brussels, Belgium, as plasmid pBT424. The strain is available from the Institut Pasteur in Paris, France.
[0115]From pBT424 the BamHl-BclI fragment containing the start codon of the delta endotoxin gene and the first 1939 bp, necessary for expression of the toxin, was cut and cloned downstream of the pTRl * promoter of plasmid pPCV520 (Koncz and Schell, 1986), cut with BamHI and Belli. The correct orientation was verified with BamHI and PstI (0.7 kb in the correct orientation) . The complete double expression cassette pTRl '-BTtox terminator gene 4/pTR2 '-nptll terminator ocs was cut again with Kpnl and partially with PstI as a 5.7 kb fragment and cloned into the corresponding restriction sites of pUCSSAcDsl9. The terminator of gene 4 contains different stop codons in the three reading frames upstream of the polyadenylation signal. The resulting plasmid (pUCSSDsl9BTKA) (Fig. 12a) was conjugated to Agrobacterium and integrated in pGV3850 by additional selection with spectinomycin as described before (Fig. 12b) .
[0116]Resistance to kanamycin could be shown after transformation to tobacco.
[0117]The construct was used for making maize lines resistant to Pyralidae, in the manner as described in Comparative Example 2, with the difference that use was made of a second Agrobacterium strain containing a transposase gene without transposon ends (pMHIOLTS) .
[0119]Construction of a recombinant plasmid containing the transposase gene of Ac under control of the CaMV 35S promoter or of the pTRl ' promoter of Agrobacterium Ach5 T-DNA.
The plasmid pMHIO (Fig. 5a) was cut with £≤£.I and the 4810bp Ac9 wx fragment, cloned into pUC19, cut wit PstI. This plasmid was 7500bp and will be referred to hereinbelow ds pUC19Ac9wx. The work was further done with the orientation 1, namely the one in which the BamHI sites are remotest from each other.
[0120]The Ac9 wx fragment was cut again, as BamHI fragment. This fragment was partially cut with CfrlOI and two fragments were stored: the 3.4 kb CfrlOI fragment and the 4.2 kb BamHI- CfrlOI fragment . The first contained a transposase gene without a promoter and the second a transposase gene with a promoter. The 4.2 kb fragment was cloned into pUCSS19, cut with Xmal and BamHI. This plasmid (8.5kb) contained the normal transposase gene of Ac but no transposon ends (pUCSS19tpnAc9PAc9) and could be transmitted by conjugation to Agrobacterium and integrated in the T-DNA of pGV3850 by additional selection of spectinomycin resistance. The 3.4 kb fragment was cloned into pUC19, cut with Xmal. and the orientation was determined using the Xbal site in the MCS and in the transposase gene (position 1260 of Ac9Wx) . This plasmid (7.7 kb) is called pUCSS19tpnAc9cds) and may serve for cloning new promoters upstream of the transposase gene.
[0121]For further use pUCSS19tpnAc9cds was taken in orientation 1, the Xbal fragment from this plasmid being 2.5 kb. This plasmid was cut with PstI and BamHI and the CaMV 35S promoter fragment from pBl221, cut with PstI and BamHI. was cloned into it .
[0122]The resulting plasmid is called pUCSS19tpnAc9P35S (Fig. 11a) . The pTRl'2' double promoter of the T-DNA of Agrobacterium Ach5 (Velten et al, 1984) was obtained as a BamHl-Pstl fragment from the plasmid pPCV520 (Koncz and Schell, 1986) and was cloned into the BamHI and PstI sites of pUCSS19tpnAc9cds. The resulting plasmid (pUCSS19tpnAc9PTRl' ) contains the transposase gene under control of the pTRl ' promoter (Fig. lib) .
[0123]Both plasmids can be conjugated, as described before, to
Agrobacterium and integrated in pGV3850 by additional selection for spectinomycin (Fig. lie) . The use of the pTRl ' promoter has the advantage that in case of icroinjection or bombardment of Agrobacterium under high pressure this promoter strongly expresses in the wounded tissue.
[0125]Construction of recombinant plasmids containing a chimeric transposon and a transposase gene without transposon ends .
[0126]Recombinants between a pMHIO derivative with a chimeric transposon and pUC19 derivatives with a chimeric transposase gene were selected in vivo in MC1061 bacteria after transformation with both plasmids and selection for ampicillin and spectinomycin, followed by alternating growth cycles with selection for ampicillin and then spectinomycin. This results in cultures in which recombinant cointegrates of pMHIO and pUC derivatives occur in 50% of the bacteria.
[0127]The constructs tpnP35S and tpnPTRl ' were cloned as Kpnl- E≤tl from their respective pUCSS plasmids into pUC19 containing the ampicillin resistance gene. The employed pMHIO derivatives were: pMH10Dsl5GUS, pMH10Ds34CAT, pMH10Dsl2IAA and pMHHDs4IPTkan and are all carriers of a spectinomycin resistance gene. The cointegrate of the recombinant plasmids ρMHHDs4IPTkan and pUCSS19Ac9P35S is shown in Fig. 13 by way of example. These cointegrate plasmids were transmitted by transformation to the strain HB101 and after selection for ampicillin and spectinomycin they were characterized by plasmid preparation and restriction enzyme analysis. Then the plasmids were transmitted by conjugation to Agrobacterium. as described before.
[0128]The resulting Agrobacterium strains are an example of the use shown in Fig. 1.
[0129]The resulting recombinant Agrobacterium strains were tested on tobacco for expression of respectively nopaline synthase (pGV3850 marker) , uidA, cat, root formation and shoot
formation and finally for transposition, before they were used on cereals .
[0131]Bolivar, F. et al., Gene 2 : 95-113 (1977) Budar, F. et al., Plant ScienceL '- 195-206 (1986) Gielen, J. et al. EMBO J. 3_: 835-846 (1984)
[0132]Hedges, R.W. and Jacob, A., Mol. Gen. Genet. 132: 31-40 (1974) Hδfte, H. et.al., Eur. J. Biochem. 161, 273-280 (1986) Inze, D. et al., Mol. Gen. Genet. 19 : 265-274 (1984)
[0133]Klier, A. et al. , EMBO J. 1, 791-799 (1982)
[0134]Koncz, C. and Schell, J., Molec. Gen. Genet. 204 :383-396 (1986)
[0135]Leemans, J. et al., J. Mol. Appl. Genet. 1:149-164 (1981) Mϋller-Neumann, M. , Yoder, I. and Starlinger, P., Mol. Gen.
[0136]Genet. 198: 19-24 (1984)
[0137]Pohlman, R.F. et al. , Cell 27: 635-643 (1984)
[0138]Van Haute, E. et al., EMBO J. 2: 411-418 (1983)
[0139]Velten, J. et al., EMBO J. 3_: 2723-2730 (1984) Vernade, D. et al., J. Bacteriol. H£: 5822-5829 (1988)
[0140]Vieira, J. and Messing, J., Gene j£: 259-268 (1982)
[0141]Yanisch-Perron, C, Vieira, J. and Messing, J., Gene 22.'* 103-
[0143]Zambryski, P. et al., EMBO J. 2 : 2143-2150 (1983)