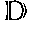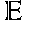CROSS-REFERENCE TO RELATED APPLICATIONS
[0001]This application claims benefit of U.S. Provisional Application No. 60/676,604, filed Apr. 29, 2005, which is hereby incorporated herein by reference in its entirety.
BACKGROUND OF THE INVENTION
[0002]Adeno-associated virus (AAV) is a member of the Parvoviridae, a virus family characterized by a single stranded linear DNA genome and a small icosahedral shaped capsid measuring about 20 nm in diameter. AAV was first described as a contamination of tissue culture grown simian virus 15, a simian adenovirus and was found dependent on adenovirus for measurable replication. This lead to its name, adeno-associated virus, and its classification in the genus Dependovirus (reviewed in Hoggan et al., 1970). AAV is a common contaminant of adenovirus samples and has been isolated from human virus samples (AAV2, AAV3, AAV5), from samples of simian virus-15 infected cells (AAV1, AAV4) as well as from stocks of avian (AAAV) (Bossis and Chiorini, 2003), bovine, canine and ovine adenovirus and laboratory adenovirus type 5 stock (AAV6). DNA spanning the entire rep-cap ORFs of AAV7 and AAV8 was amplified by PCR from heart tissue of rhesus monkeys (Gao et al., 2002). With the exception of AAVs 1 and 6, all cloned AAV isolates appear to be serologically distinct. Nine isolates have been cloned, and recombinant viral stocks have been generated from each isolated virus.
[0003]AAV appears to commonly infect humans. 50%-80% of adults in North America are seropositive for AAV. A steep rise in antibody response against AAV 1-3 was observed in the age group between 1-10 years (Blacklow et al., 1968). AAV 2 and 3 were readily isolated from anal and throat specimens from children (Blacklow et al., 1967) whereas isolation from adults was not observed. It appears that AAV spreads primarily in the young population (Hoggan, 1970). Prevalence of antibodies against AAV was found to be similar in Europe, Brazil and Japan, which suggests a global spread of AAV (Erles et al., 1999). Infection with AAV appears to be benign in man and laboratory animals. Currently, no disease has been associated with AAV infections.
[0004]AAV2 is the best characterized adeno-associated virus and will be discussed as an AAV prototype. The AAV2 genome consists of a linear single stranded DNA of 4,780 nucleotides. Both polarities of DNA are encapsulated by AAV with equal efficiency. The AAV2 genome contains 2 open reading frames (ORF) named rep and cap. The rep ORF encodes the non-structural proteins that are essential for viral DNA replication, packaging and AAV integration. The cap ORF encodes the capsid proteins. The rep ORF is transcribed from promoters at map units P5 and P19. The rep transcripts contain an intron close to the 3′ end of the rep ORF and can be alternatively spliced. The rep ORF is therefore expressed as 4 partially overlapping proteins, which were termed according to their molecular weight Rep78, 68, 52 and 40. The cap ORF is expressed from a single promoter at P40. By alternative splicing and utilization of an alternative ACG start codon, cap is expressed into the capsid proteins VP1-3 which range in size from 65-86 kDa. VP3 is the most abundant capsid protein and constitutes 80% of the AAV2 capsid. All viral transcripts terminate at a polyA signal at map unit 96.
[0005]During a productive AAV2 infection, unspliced mRNAs from the p5 promoter encoding Rep78 are the first detectable viral transcripts. In the course of infection, expression from P5, P19 and P40 increase to 1:3:18 levels respectively. The levels of spliced transcripts increased to 50% for P5, P19 products and 90% of P40 expressed RNA (Mouw and Pintel, 2000).
[0006]The AAV2 genome is terminated on both sides by inverted terminal repeats (ITRs) of 145 nucleotides (nt). 125 nt of the ITR constitute a palindrome which contains 2 internal palindromes of 21 nt each. The ITR can fold back on itself to generate a T-shaped hairpin with only 7 non-paired bases. The stem of the ITR contains a Rep binding site (RBS) and a sequence that is site and strand specifically cleaved by Rep—the terminal resolution site (TRS). The ITR is essential for AAV2 genome replication, integration and contains the packaging signals.
[0007]The single-stranded AAV2 genome is packaged into a non-enveloped icosahedral shaped capsid of about 20-25 nm diameter. The virion consists of 26% DNA and 74% protein and has a density of 1.41 g/cm3. AAV2 particles are extremely stable and can withstand heating to 60° C. for 1 hour, extreme ph, and extraction with organic solvents.
[0008]Rep proteins are involved in almost every step of AAV2 replication including AAV2 genome replication, integration, and packaging. Rep78 and Rep68 possess ATPase, 3′-5′ helicase, ligase and nicking activities and bind specifically to DNA. Rep52 and Rep40 appear to be involved in the encapsidation process and encode ATPase and 3′-5′ helicase activities. Mutational analysis suggests a domain structure for Rep78. The N-terminal 225 aa are involved in DNA binding, DNA nicking and ligation. Rep78 and Rep68 recognize a GCTC repeat motif in the ITR as well as in a linear truncated form of the ITR (Chiorini et al., 1994) with similar efficiencies. Rep78 and Rep68 possess a sequence and strand specific endonuclease activity, which cleaves the ITR at the terminal resolution site (TRS). Rep endonuclease activity is dependent on nucleoside triphosphate hydrolysis and presence of metal cations. Rep 78 and 68 can also bind and cleave single stranded DNA in a NTP independent matter. In addition, Rep78 catalyzes rejoining of single stranded DNA substrates originating from the AAV2 origin of replication—i.e., sequences containing a rep binding and terminal resolution element.
[0009]The central region of AAV2 Rep78, which represents the N-terminus of Rep52 and Rep40, contains the ATPase and helicase activities as well as nuclear localization signals. The helicase activity unwinds DNA-DNA and DNA-RNA duplexes, but not RNA-RNA. The ATPase activity is constitutive and independent of a DNA substrate. The C-terminus of Rep78 contains a potential zinc-finger domain and can inhibit the cellular serine/threonine kinase activity of PKA as well as its homolog PRKX by pseudosubstrate inhibition. Rep68 which is translated from a spliced mRNA that encodes the N-terminal 529 amino acids (aa) of Rep78 fused to 7 aa unique for Rep68, doesn't inhibit either PKA or PRKX. In addition to these biochemical activities, Rep can affect intracellular conditions by protein-protein interactions. Rep78 binds to a variety of cellular proteins including transcription factors like SP-1, high-mobility-group non-histone protein 1 (HMG-1) and the oncosuppressor p53. Overexpression of Rep results in pleiotrophic effects. Rep78 disrupts cell cycle progression and inhibits transformation by cellular and viral oncogenes. In susceptible cell lines, overexpression of Rep resulted in apoptosis and cell death. Several of Rep78 activities contribute to cytotoxicity, including its constitutive ATPase activity, interference with cellular gene expression and protein interactions.
[0010]The first step of an AAV infection is binding to the cell surface. Receptors and coreceptors for AAV2 include heparan sulfate proteoglycan, fibroblast growth factor receptor-1, and αvβ5integrins whereas N-linked 2,3-linked sialic acid is required for AAV5 binding and transduction (Walters et al., 2001). In HeLa cells, fluorescently labeled AAV2 particles appear to enter the cell via receptor-mediated endocytosis in clathrin coated pits. More than 60% of bound virus was internalized within 10 min after infection. Labeled AAV particles are observed to have escaped from the endosome, been trafficked via the cytoplasm to the cell nucleus and accumulated perinuclear, before entering the nucleus, probably via nuclear pore complex (NPC). AAV2 particles have been detected in the nucleus, suggesting that uncoating takes place in the nucleus (Bartlett et al., 2000; Sanlioglu et al., 2000). AAV5 is internalized in HeLa cells predominantly by clathrin coated vesicles, but to a lesser degree also in noncoated pits. AAV particles can also be trafficked intercellularly via the Golgi apparatus (Bantel-Schaal et al., 2002). At least partial uncoating of AAV5 was suggested to take place before entering the nucleus since intact AAV5 particles could not be detected in the nucleus (Bantel-Schaal et al., 2002) After uncoating, the single stranded genome is converted into duplex DNA either by leading strand synthesis or annealing of input DNA of opposite polarity. AAV replication takes place within the nucleus.
[0011]During a co-infection with a helper virus such as Adenovirus, herpes simplex virus or cytomegalovirus, AAV is capable of an efficient productive replication. The helper functions provided by Adenovirus have been studied in great detail. In human embryonic kidney 293 cells, which constitutively express the Adenovirus E1A and E1B genes, the early Adenovirus gene products of E2A, E4 and VA were found sufficient to allow replication of recombinant AAV. Allen et al. reported that efficient production of rAAV is possible in 293 cells transfected with only an E4orf6 expression plasmid (Allen et al., 2000). E1A stimulates S phase entry and induces unscheduled DNA synthesis by inactivating the pRB checkpoint at the G1/S border by interaction with pRB family proteins which results in the release of E2F (reviewed in (Ben-Israel and Kleinberger, 2002). This leads to either induction or activation of enzymes involved in nucleotide synthesis and DNA replication. Since unscheduled DNA synthesis is a strong apoptotic signal, anti-apoptotic functions are required. E1B-19k is a Bcl-2 homolog and E1B-55k is a p53 antagonist. Both proteins have anti-apoptotic functions. E4orf6 forms a complex with E1B-55k and results in degradation of p53. It is also reported to cause S-phase arrest (Ben-Israel and Kleinberger, 2002). E2A encodes a single strand DNA binding protein, which appears to be non-essential for DNA replication but effects gene expression (Chang and Shenk, 1990) (Fields 39, 40). The VA transcription unit affects AAV2 RNA stability and translation (Janik et al., 1989). E1A has a more direct effect on AAV2 gene expression. The cellular transcription factor YY-1 binds and inhibits the viral P5 promoter. E1A relieves this transcriptional block. None of the late Ad gene products have been found to be essential for AAV2 replication. The main function of the helper virus appears to be the generation of a cellular environment with active DNA replication machinery and blocked pro-apoptotic functions that allows high-level AAV replication rather than a direct involvement in AAV replication.
[0012]While AAV is usually dependent on a helper virus for efficient replication, low level AAV replication was observed under conditions of genotoxic stress (Yakinoglu et al., 1988; Yakobson et al., 1989). AAV DNA replication and particle formation was also observed in differentiating keratinocytes in the absence of helper virus infection (Meyers et al., 2000). This demonstrates that AAV is not defective per se but rather depends on the helper virus to establish the favorable cellular condition and to provide factors for efficient replication
[0013]The ability of AAV vectors to infect dividing and non-dividing cells, establish long-term transgene expression, and the lack of pathogenicity has made them attractive for use in gene therapy applications. Lack of cross competition in binding experiments suggests that each AAV serotype may have a distinct mechanism of cell entry. Comparison of the cap ORFs from different serotypes has identified blocks of conserved and divergent sequence, with most of the latter residing on the exterior of the virion, thus explaining the altered tissue tropism among serotypes (19-21, 48, 56). Vectors based on new AAV serotypes may have different host range and different immunological properties, thus allowing for more efficient transduction in certain cell types. In addition, characterization of new serotypes will aid in identifying viral elements required for altered tissue tropism.
BRIEF SUMMARY OF THE INVENTION
[0014]Provided herein are adeno-associated viruses (AAV) and vectors derived therefrom. Thus, the present invention relates to AAV vectors for and methods of delivering nucleic acids to cells of subjects.
[0015]Additional advantages of the disclosed method and compositions will be set forth in part in the description which follows, and in part will be understood from the description, or may be learned by practice of the disclosed method and compositions. The advantages of the disclosed method and compositions will be realized and attained by means of the elements and combinations particularly pointed out in the appended claims. It is to be understood that both the foregoing general description and the following detailed description are exemplary and explanatory only and are not restrictive of the invention as claimed.
BRIEF DESCRIPTION OF THE DRAWINGS
[0016]The accompanying drawings, which are incorporated in and constitute a part of this specification, illustrate several embodiments of the disclosed method and compositions and together with the description, serve to explain the principles of the disclosed method and compositions.
[0017]FIG. 1 shows evolutionary relationship among human, non-human primate AAVs, and AAV contaminants detected in adenovirus stocks. The phylogenetic tree is based on merged ClustalW alignments of VP1 sequences. VR numbers are identifiers of the ATCC virus collection.
[0018]FIG. 2 shows rAAV6, rAAV(VR-195), and rAAV(VR-355) are neutralized by pooled human IgGs. COS cells were transduced with a pre-incubation mixture consisting of rAAV-6, rAAV(VR-195), and rAAV(VR-355) expressing GFP and human IgGs at the indicated concentrations. Twenty-four hours post-inoculation, transduction was analyzed by flow cytometry and graphed as percent transduction of the untreated control. Values are means from three experiments; error bars represent standard deviations.
[0019]FIG. 3 shows inhibitory effect of heparin on of COS cell transduction. COS cells were transduced with a pre-incubation mixture consisting out of rAAV2, rAAV5, rAAV6, rAAV(VR-195), and rAAV(VR-355) expressing GFP and heparin at the indicated concentrations. 24 h post-inoculation, transduction was analyzed by flow cytometry. Values are means from three experiments; error bars represent standard deviations.
[0020]FIG. 4 shows charge-dependency of AAV6, rAAV(VR-195), and rAAV(VR-355) transduction of COS cells. COS cells were transduced with rAAV6, rAAV(VR-195), and rAAV(VR-355) in medium containing the indicated concentrations of NaCl. Twenty-four hour postinoculation, transduction was analyzed by flow cytometry. Values are means from three experiments; error bars represent standard deviations
[0021]FIG. 5 shows neuraminidase treatment blocks rAAV6, rAAV(VR-195), and rAAV(VR-355) transduction and cell binding. FIG. 5A shows gene transfer mediated by rAAV6, rAAV(VR-195), and rAAV(VR-355) expressing GFP in COS cells, following neuraminidase pretreatment. FIG. 5B shows binding. Values are means from three experiments; error bars represent standard deviations
[0022]FIG. 6 shows effects of lectins on rAAV6, rAAV(VR-195), and rAAV(VR-355) transduction of COS cells. After pre-incubation with ECL, LCA, MalII or STL, COS cells were transduced with rAAV-6, rAAV(VR-195), and rAAV(VR-355) expressing GFP in the presence of the indicated lectin. Twenty-four hour post-inoculation, transduction was analyzed by flow cytometry. Values are means from two experiments done in duplicate; error bars represent standard deviations.
[0023]FIG. 7 shows transduction efficiency in human cancer cell lines. The indicated cell lines were transduced with 1×108particles of rAAV6, rAAV(VR-195), and rAAV(VR-355) expressing a nuclear localized GFP. Twenty-four hour post-inoculation, transduction was analyzed by flow cytometry. Values are means from three experiments; error bars represent standard deviations. * COS cells transduction is given as transducing units/2×107particles.
[0024]FIG. 8 shows AAV6 competition. COS cells were transduced with a constant amount of rAAV6, rAAV(VR-195) or rAAV(VR-355) expressing GFP after 60 min pre-incubation with increasing titers of rAAV6-lacZ. Forty-eight hour post-inoculation, transduction was analyzed by flow cytometry. Values are means from three experiments; error bars represent standard deviations.
[0025]FIG. 9 shows effects of sugars on rAAV mediated transduction. Cos cells were transduced with GFP encoding rAAVs that were pre-incubated with various sugar monomers and polymers. 24 h post-inoculation, transduction was analyzed by flow cytometry. Heparin is the attachment factor for AAV2 that mediates initial binding of AAV2 to the cell. Extracellular heparin binds to the virus and blocks the attachment of the virus to the cell. None of the sugars tested blocked transduction of AAV-X1, AAV-X5, AAV-X25 or AAV-X26.
[0026]FIG. 10 shows effects of lectins on rAAV transduction of COS cells. After pre-incubation with various lectins, COS cells were transduced with rAAV-6, AAV-X1 and AAV-X25 expressing GFP in the presence of the indicated lectin. 24 h post-inoculation, transduction was analyzed by flow cytometry. All AAVs have a distinct profile on the lectin panel.
[0027]FIG. 11 shows transduction efficiency of rAAVs in cell lines. Cells were transduced with various rAAV serotypes encoding GFP. Two days after inoculation, cells were analyzed for GFP expression by flow cytometry. The transduction profiles of AAV-X1, AAV-X5, AAV-X25 and AAV-X26 were different from that of known AAVs.
[0028]FIG. 12 shows rAAV12 COS cell transduction is not inhibited by heparin. COS cells were transduced with a pre-incubation mixture consisting out of rAAV2-GFP, or rAAV12-GFP expressing GFP and heparin at the indicated concentrations. 24 h post-inoculation, transduction was analyzed by flow cytometry. Values are means from three experiments; error bars represent standard deviations.
[0029]FIG. 13 shows rAAV12 transduction is independent of cell surface sialic acid. COS cells were pretreated with the V. cholera neuraminidase to remove exposed sialic acids groups before the cells were transduced with rAAV2, rAAV4, rAAV5 and rAAV12-GFP. Gene transfer was determined by flow cytometry. Values are means from three experiments; error bars represent standard deviations.
[0030]FIG. 14 shows rAAV12 transduction is protease sensitive and does not require glycosphingolipids. COS cells were proteolytically digested with trypsin (A) or treated with the glycosphingolipids synthesis inhibitors PPMP (B) prior to transduction with rAAV2-GFP, rAAV12-GFP and rBAAV-GFP. Gene transfer in these cells was compared to untreated cultures. Values are means from three experiments; error bars represent standard deviations.
[0031]FIG. 15 shows extracellular mannosamine inhibits rAAV12 transduction. COS cells were transduced with a pre-incubation mixture of rAAV2-GFP, or rAAV12-GFP and mannosamine at the indicated concentrations. 24 h post-inoculation, transduction was analyzed by flow cytometry. Values are means from three experiments; error bars represent standard deviations.
[0032]FIG. 16 shows rAAV12 has a broad tropism. Transduction efficiency of rAAV12 was compared to rAAV4 in 13 human cancer cell lines. Cells were transduced with particles of either rAAV12-GFP or rAAV4-GFP. Transduction was analyzed by flow cytometry 28 h after virus inoculation. Values are means from three experiments; error bars represent standard deviations.
[0033]FIG. 17 shows rAAV12 is highly resistant to neutralization by human IgGs. rAAV2-GFP, rAAV6-GFP and rAAV12-GFP were incubated with pooled human IgGs prior to transduction of COS cells. 24 h post-inoculation, transduction was analyzed by flow cytometry. Transduction efficiencies relative to an untreated control were plotted. Values are means from three experiments; error bars represent standard deviations.
[0034]FIG. 18 shows rAAV12 transduces salivary glands and skeletal muscles in vivo. Male Balb/c mice were administered 109particlesof either AAV2-hEPO or AAV12-hEPO by retrograde ductal delivery to both submandibular glands or in both their tibialis anterior (two injection sites per muscle). Secretion of hEPO in mouse serum was determined 4 weeks after transduction by an ELISA test. Error bars represent standard deviations.
DETAILED DESCRIPTION OF THE INVENTION
[0035]The disclosed method and compositions may be understood more readily by reference to the following detailed description of particular embodiments and the Example included therein and to the Figures and their previous and following description.
[0036]It is to be understood that the disclosed methods and compositions are not limited to specific synthetic methods, specific analytical techniques, or to particular reagents unless otherwise specified, and, as such, may vary. It is also to be understood that the terminology used herein is for the purpose of describing particular embodiments only and is not intended to be limiting.
[0037]Disclosed are materials, compositions, and components that can be used for, can be used in conjunction with, can be used in preparation for, or are products of the disclosed method and compositions. These and other materials are disclosed herein, and it is understood that when combinations, subsets, interactions, groups, etc. of these materials are disclosed that while specific reference of each various individual and collective combinations and permutation of these compounds may not be explicitly disclosed, each is specifically contemplated and described herein. For example, if a vector is disclosed and discussed and a number of modifications that can be made to a number of molecules including the promoters and ITRs are discussed, then each and every combination and permutation of the promoters and ITRs, and the modifications that are possible, are specifically contemplated unless specifically indicated to the contrary. Thus, if a class of molecules A, B, and C are disclosed as well as a class of molecules D, E, and F and an example of a combination molecule, A-D is disclosed, then even if each is not individually recited, each is individually and collectively contemplated. Thus, is this example, each of the combinations A-E, A-F, B-D, B-E, B-F, C-D, C-E, and C-F are specifically contemplated and should be considered disclosed from disclosure of A, B, and C; D, E, and F; and the example combination A-D. Likewise, any subset or combination of these is also specifically contemplated and disclosed. Thus, for example, the sub-group of A-E, B-F, and C-E are specifically contemplated and should be considered disclosed from disclosure of A, B, and C; D, E, and F; and the example combination A-D. This concept applies to all aspects of this application including, but not limited to, steps in methods of making and using the disclosed compositions. Thus, if there are a variety of additional steps that can be performed it is understood that each of these additional steps can be performed with any specific embodiment or combination of embodiments of the disclosed methods, and that each such combination is specifically contemplated and should be considered disclosed.
[0038]It must be noted that as used herein and in the appended claims, the singular forms “a”, “an”, and “the” include plural reference unless the context clearly dictates otherwise. Thus, for example, reference to “a vector” includes a plurality of such vectors, reference to “the vector” is a reference to one or more vectors and equivalents thereof known to those skilled in the art, and so forth.
[0039]“Optional” or “optionally” means that the subsequently described event, circumstance, or material may or may not occur or be present, and that the description includes instances where the event, circumstance, or material occurs or is present and instances where it does not occur or is not present.
[0040]Ranges may be expressed herein as from “about” one particular value, and/or to “about” another particular value. When such a range is expressed, also specifically contemplated and considered disclosed is the range from the one particular value and/or to the other particular value unless the context specifically indicates otherwise. Similarly, when values are expressed as approximations, by use of the antecedent “about,” it will be understood that the particular value forms another, specifically contemplated embodiment that should be considered disclosed unless the context specifically indicates otherwise. It will be further understood that the endpoints of each of the ranges are significant both in relation to the other endpoint, and independently of the other endpoint unless the context specifically indicates otherwise. Finally, it should be understood that all of the individual values and sub-ranges of values contained within an explicitly disclosed range are also specifically contemplated and should be considered disclosed unless the context specifically indicates otherwise. The foregoing applies regardless of whether in particular cases some or all of these embodiments are explicitly disclosed.
[0041]Unless defined otherwise, all technical and scientific terms used herein have the same meanings as commonly understood by one of skill in the art to which the disclosed method and compositions belong. Although any methods and materials similar or equivalent to those described herein can be used in the practice or testing of the present method and compositions, the particularly useful methods, devices, and materials are as described. Publications cited herein and the material for which they are cited are hereby specifically incorporated by reference. Nothing herein is to be construed as an admission that the present invention is not entitled to antedate such disclosure by virtue of prior invention. No admission is made that any reference constitutes prior art. The discussion of references states what their authors assert, and applicants reserve the right to challenge the accuracy and pertinency of the cited documents. It will be clearly understood that, although a number of publications are referred to herein, such reference does not constitute an admission that any of these documents forms part of the common general knowledge in the art.
[0042]Throughout the description and claims of this specification, the word “comprise” and variations of the word, such as “comprising” and “comprises,” means “including but not limited to,” and is not intended to exclude, for example, other additives, components, integers or steps.
[0043]Those skilled in the art will recognize, or be able to ascertain using no more than routine experimentation, many equivalents to the specific embodiments of the method and compositions described herein. Such equivalents are intended to be encompassed by the following claims.
[0044]Provided herein are new recombinant adeno-associated viruses (AAVs) designated AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, and AAV-X26. The term AAVX is used herein to refer generally to the new AAVs disclosed herein. Thus, AAVX can refer to one or more or all of AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, or AAV-X26. The application provides the isolation, subcloning, and sequencing of the disclosed AAVXs. These viruses have one or more of the characteristics described below. In one embodiment, the compositions provided herein do not include wild-type AAV. The methods provided herein can use either wild-type AAV or recombinant AAV-based delivery. Thus, in one embodiment, the methods provided herein do not use wild-type AAV.
[0045]Provided herein are recombinant AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, and AAV-X26 particles, recombinant AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, and AAV-X26 vectors and recombinant AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, and AAV-X26 virions. As used herein, “recombinant” refers to nucleic acids, vectors, polypeptides, or proteins that have been generated using DNA recombination (cloning) methods and are distinguishable from native or wild-type nucleic acids, vectors, polypeptides, or proteins. An AAVX particle is a viral particle comprising an AAVX capsid protein. A recombinant AAVX vector is a nucleic acid construct that comprises at least one unique, isolated nucleic acid of AAVX. The recombinant AAVX vector can further comprise at least one non-AAVX nucleic acid. As used herein, a “virion” refers to an infectious virus particle, and “infectious” refers to the ability of a virion to deliver genetic material to a cell. Thus, a recombinant AAVX virion is a particle containing a recombinant AAVX vector, wherein the particle can be either an AAVX particle as described herein or a non-AAVX particle. Alternatively, a recombinant AAVX virion can be an AAVX particle containing a recombinant vector, wherein the vector can be either an AAVX vector as described herein or a non-AAVX vector. An AAVX particle can further be an “empty particle”, wherein the particle does not contain a nucleic acid, vector or plasmid, and is therefore not infectious. These vectors, particles, virions, nucleic acids and polypeptides are described below.
[0046]Provided herein are nucleotide sequences of AAVX genomes and vectors and particles derived therefrom. Specifically provided is an AAVX nucleic acid vector. Thus, provided is a nucleic acid vector, comprising an AAVX-specific nucleic acid or a nucleic acid encoding an AAVX-specific protein. The AAVX-specific nucleic acid can be a pair of AAVX inverted terminal repeats (ITRs) or an AAVX promoter. The nucleic acid encoding an AAVX-specific protein can be an AAVX capsid protein or an AAVX Rep protein. Thus, the provided AAVX nucleic acid vector need only have an AAVX ITR, an AAVX promoter, an AAVX Rep or an AAVX capsid to be an AAVX nucleic acid vector.
[0047]The AAV ITR functions as an origin of replication for packaging of recombinant AAV particles. The minimum sequence necessary for this activity is the TRS site where Rep cleaves in order to replicate the virus. Minor modifications in an ITR are contemplated and are those that will not interfere with the hairpin structure formed by the ITR as described herein and known in the art. Furthermore, to be considered within the term it must retain the Rep binding site described herein. For example, the D− region of the AAV2 ITR, a single stranded region of the ITR (the D+ region is the reverse complement of the D− region), inboard of the TRS site, has been shown to bind a factor, which, depending on its phosphorylation state, correlates with the conversion of the AAV2 from a single stranded genome to a transcriptionally active form that allows for expression of the viral DNA. For example, this region is conserved between AAV2, AAV3, AAV4, and AAV6 but is divergent in AAV5 and BAAV. Further, as disclosed herein, the TRS signal (e.g., aa 176-181 of SEQ ID NO:40) and Rep Binding site (e.g., aa 195-210 of SEQ ID NO:40) is conserved between AAV2 and AAV-X26.
[0048]In one aspect, the AAVX nucleic acid vector comprises an AAVX ITR. Thus, the AAVX nucleic acid vector can comprise an AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, or AAV-X26 ITR. In another aspect, the AAVX nucleic acid vector comprises an ITR from any AAV serotype. Thus, the AAVX nucleic acid vector can comprise an AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, or BAAV ITR.
[0049]In one aspect, the AAVX nucleic acid vector comprises an AAVX promoter. Thus, the promoter can be an AAVX p5, p19 or p40 promoter. Thus, the promoter can be an AAV-X1 p5 promoter, an AAV-X1b p5 promoter, an AAV-X5 p5 promoter, an AAV-X19 p5 promoter, n AAV-X21 p5 promoter, an AAV-X22 p5 promoter, an AAV-X23 p5 promoter, an AAV-X24 p5 promoter, an AAV-X25 p5 promoter, or an AAV-X26 p5 promoter.
[0050]In another aspect, the promoter can be a promoter from any of the AAV serotypes. Thus, the promoter can be an AAV1 p5 promoter, an AAV2 p5 promoter, an AAV3 p5 promoter, an AAV4 p5 promoter, AAV5 p5 promoter, an AAV6 p5 promoter, an AAAV p5 promoter, a BAAV p5 promoter, or an AAVX p5 promoter.
[0051]Furthermore, smaller fragments of an AAV p5 promoter that retain promoter activity can readily be determined by standard procedures including, for example, constructing a series of deletions in the p5 promoter, linking the deletion to a reporter gene, and determining whether the reporter gene is expressed, i.e., transcribed and/or translated.
[0052]In yet another aspect, the promoter of the AAVX nucleic acid vector can be any desired promoter, selected by known considerations, such as the level of expression of a nucleic acid functionally linked to the promoter and the cell type in which the vector is to be used. That is, the promoter can be tissue/cell-specific. Promoters can be prokaryotic, eukaryotic, fungal, nuclear, mitochondrial, viral or plant promoters. Promoters can be exogenous or endogenous to the cell type being transduced by the vector. Promoters can include, for example, bacterial promoters, or known strong promoters such as SV40 or the inducible metallothionein promoter. Additionally, chimeric regulatory promoters for targeted gene expression can be utilized. Examples of these regulatory systems, which are known in the art, include the tetracycline based regulatory system which utilizes the tet transactivator protein (tTA), a chimeric protein containing the VP16 activation domain fused to the tet repressor of Escherichia coli, the IPTG based regulatory system, the CID based regulatory system, and the Ecdysone based regulatory system (No, D., et al., Proc Natl Acad Sci USA. 93(8):3346-3351 (1996)). Other promoters include promoters derived from actin genes, immunoglobulin genes, cytomegalovirus (CMV), adenovirus, bovine papilloma virus, adenoviral promoters, such as the adenoviral major late promoter, an inducible heat shock promoter, respiratory syncytial virus, Rous sarcomas virus (RSV), etc.
[0053]In one aspect, the AAVX nucleic acid vector comprises a nucleic acid encoding an AAVX Rep protein. Thus, the Rep protein can be an AAV-X1 Rep protein, an AAV-X1b Rep protein, an AAV-X5 Rep protein, an AAV-X19 Rep protein, n AAV-X21 Rep protein, an AAV-X22 Rep protein, an AAV-X23 Rep protein, an AAV-X24 Rep protein, an AAV-X25 Rep protein, or an AAV-X26 Rep protein. In another aspect, the AAVX nucleic acid vector comprises a nucleic acid encoding a Rep protein from any AAV serotype. Thus, the AAVX nucleic acid vector can comprise a nucleic acid encoding an AAV1 Rep protein, an AAV2 Rep protein, an AAV3 Rep protein, an AAV4 Rep protein, an AAV5 Rep protein, an AAV6 Rep protein, an AAV7 Rep protein, an AAV8 Rep protein, an AAV9 Rep protein, an AAV10 Rep protein, an AAV11 Rep protein, an AAAV Rep protein, or an BAAV Rep protein. For all AAV serotypes, the AAV Rep proteins can be selected from a group consisting of Rep78, Rep68, Rep52 and Rep40.
[0054]The AAV-X1 Rep protein of an AAV-X1 nucleic acid vector can be encoded by a nucleic acid sequence comprising nucleotides 1-743, 744, 745, 746, 747, 748, 749, 750, 751, 752, 752, 754, 755, 756, 757, 758, 759, 760, 761, 762, or 763 of SEQ ID NO:1. The AAV-X1 Rep protein can be encoded by a nucleic acid sequence of SEQ ID NO:48. The AAV-X1 Rep protein can comprise the amino acid sequence of SEQ ID NO:49. The AAV-X1b Rep protein of an AAV-X1b nucleic acid vector can be encoded by a nucleic acid sequence comprising nucleotides 1-2016, 2017, 2018, 2019; 2020, 2021, 2022, 23 2024, 2025, 2026, 2027, 2028, 2029, 2030, 2031, 2032, 2033, 2034, 2035, or 2036 of SEQ ID NO:2. The AAV-X5 Rep protein of an AAV-X5 nucleic acid vector can be encoded by a nucleic acid sequence comprising nucleotides 1-1926, 1927, 1928, 1929, 1930, 1931, 1932, 1933, 1934, 1935, 1936, 1937, 1938, 1939, 1940, 1941, 1942, 1943, 1944, 1945, or 1946 of SEQ ID NO3. The AAV-X19 Rep protein of an AAV-X19 nucleic acid vector can be encoded by a nucleic acid sequence comprising nucleotides 1-743, 744, 745, 746, 747, 748, 749, 750, 751, 752, 752, 754, 755, 756, 757, 758, 759, 760, 761, 762, or 763 of SEQ ID NO:4. The AAV-X21 Rep protein of an AAV-X21 nucleic acid vector can be encoded by a nucleic acid sequence comprising nucleotides 1-752, 753, 754, 755, 756, 757, 758, 759.760, 761, 762, 763, 764, 765, 766, 767, 768, 769, 770, 771, or 772 of SEQ ID NO:5. The AAV-X22 Rep protein of an AAV-X22 nucleic acid vector can be encoded by a nucleic acid sequence comprising nucleotides 1-748, 749, 750, 751, 752, 753, 754, 755, 756, 757, 758, 759, 760, 761, 762, 763, 764, 765, 766, 767, or 768 of SEQ ID NO:6. The AAV-X23 Rep protein of an AAV-X23 nucleic acid vector can be encoded by a nucleic acid sequence comprising nucleotides 1-746, 747, 748, 749, 750, 751, 752, 753, 754, 755, 756, 757, 758, 759, 760, 761, 762, 763, 764, 765, or 766 of SEQ ID NO:7. The AAV-X24 Rep protein of an AAV-X24 nucleic acid vector can be encoded by a nucleic acid sequence comprising nucleotides 1-752, 753, 754, 755, 756, 757, 758, 759, 760, 761, 762, 763, 764, 765, 766, 767, 768, 769, 770, 771, or 772 of SEQ ID NO:8. The AAV-X25 Rep protein of an AAV-X25 nucleic acid vector can be encoded by the a nucleic acid sequence comprising nucleotides 1-974, 975, 976, 977, 978, 979, 980, 981, 982, 983, 984, 985, 986, 987, 988, 989, 990, 991, 992, 993, or 994 of SEQ ID NO:9. The AAV-X25 Rep protein can be encoded by a nucleic acid sequence of SEQ ID NO:50. The AAV-X25 Rep protein can comprise the amino acid sequence of SEQ ID NO:51. The AAV-X26 Rep protein of an AAV-X26 nucleic acid vector can be encoded by the a nucleic acid sequence comprising nucleotides 1-789, 790, 791, 792, 793, 794, 795, 796, 797, 798, 799, 800, 801, 802, 803, 804, 805, 806, 807, 808, or 809 of SEQ ID NO:10. The AAV-X26 Rep protein can be encoded by a nucleic acid sequence of SEQ ID NO:52. The AAV-X26 Rep protein can comprise the amino acid sequence of SEQ ID NO:53.
[0055]In one aspect, the AAVX nucleic acid vector comprises a nucleic acid encoding an AAVX capsid protein. Thus, the capsid protein can be an AAV-X1 capsid protein, an AAV-X1b capsid protein, an AAV-X5 capsid protein, an AAV-X19 capsid protein, n AAV-X21 capsid protein, an AAV-X22 capsid protein, an AAV-X23 capsid protein, an AAV-X24 capsid protein, an AAV-X25 capsid protein, or an AAV-X26 capsid protein. In another aspect, the AAVX nucleic acid vector comprises a nucleic acid encoding a capsid protein from any AAV serotype. Thus, the AAVX nucleic acid vector can comprise a nucleic acid encoding an AAV 1 capsid protein, an AAV2 capsid protein, an AAV3 capsid protein, an AAV4 capsid protein, an AAV5 capsid protein, an AAV6 capsid protein, an AAV7 capsid protein, an AAV8 capsid protein, an AAV9 capsid protein, an AAV10 capsid protein, an AAV 11 capsid protein, an AAAV capsid protein, or an BAAV capsid protein. For all AAV serotypes, the AAV capsid proteins can be selected from a group consisting of VP1, VP2 and VP3.
[0056]As an example, the AAV-X1 VP1 capsid protein of an AAV-X1 particle can have the amino acid sequence of SEQ ID NO:21. The AAV-X1b VP1 capsid protein of an AAV-X1b particle can have the amino acid sequence of SEQ ID NOS:22. The AAV-X5 VP1 capsid protein of an AAV-X5 particle can have the amino acid sequence of SEQ ID NO:23. The AAV-X19 VP1 capsid protein of an AAV-X19 particle can have the amino acid sequence of SEQ ID NO:24. The AAV-X21 VP1 capsid protein of an AAV-X21 particle can have the amino acid sequence of SEQ ID NO:25. The AAV-X22 VP1 capsid protein of an AAV-X22 particle can have the amino acid sequence of SEQ ID NO:26. The AAV-X23 VP1 capsid protein of an AAV-X23 particle can have the amino acid sequence of SEQ ID NO:27. The AAV-X24 VP1 capsid protein of an AAV-X24 particle can have the amino acid sequence of SEQ ID NO:28. The AAV-X25 capsid protein of an AAV-X25 particle can have the amino acid sequence of SEQ ID NO:29. The AAV-X26 capsid protein of an AAV-X26 particle can have the amino acid sequence of SEQ ID NO:30.
[0057]The AAV-X1 VP1 capsid protein of an AAV-X1 particle can be encoded by the nucleic acid sequence of SEQ ID NO:11. The AAV-X1b VP1 capsid protein of an AAV-X1b particle can be encoded by the nucleic acid sequence of SEQ ID NO:12. The AAV-X5 VP1 capsid protein of an AAV-X5 particle can be encoded by the nucleic acid sequence of SEQ ID NO:13. The AAV-X19 VP1 capsid protein of an AAV-X19 particle can be encoded by the nucleic acid sequence of SEQ ID NO:14. The AAV-X21 VP1 capsid protein of an AAV-X21 particle can be encoded by the nucleic acid sequence of SEQ ID NO:15. The AAV-X22 VP1 capsid protein of an AAV-X22 particle can be encoded by the nucleic acid sequence of SEQ ID NO:16. The AAV-X23 VP1 capsid protein of an AAV-X23 particle can be encoded by the nucleic acid sequence of SEQ ID NOS:17. The AAV-X24 VP 1 capsid protein of an AAV-X24 particle can be encoded by the nucleic acid sequence of SEQ ID NO:18. The AAV-X25 VP1 capsid protein of an AAV-X25 particle can be encoded by the nucleic acid sequence of SEQ ID NO:19. The AAV-X26 VP1 capsid protein of an AAV-X26 particle can be encoded by the nucleic acid sequence of SEQ ID NO:20.
[0058]It should be recognized that any errors in any of the nucleotide sequences disclosed herein can be corrected, for example, by using the hybridization procedure described below with various probes derived from the described sequences such that the coding sequence can be re-isolated and re-sequenced. Rapid screening for point mutations can also be achieved with the use of polymerase chain reaction-single strand conformation polymorphism (PCR-SSCP). The corresponding amino acid sequence can then be corrected accordingly. Also, since the disclosed AAV serotypes AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, and AAV-X26 are disclosed herein to be present in defined ATCC cultures, the whole virus sequence is provided by reference to the deposit.
[0059]The AAVX-derived vector provided herein can further comprise an exogenous nucleic acid functionally linked to the promoter. By “exogenous” nucleic acid is meant any nucleic acid that is not normally found in wild-type AAVX that can be inserted into a vector for transfer into a cell, tissue or organism. The exogenous nucleic acid can be a nucleic acid not normally found in the target cell, or it can be an extra copy or copies of a nucleic acid normally found in the target cell. The terms “exogenous” and “heterologous” are used herein interchangeably.
[0060]By “functionally linked” is meant that the promoter can promote expression of the exogenous nucleic acid, as is known in the art, and can include the appropriate orientation of the promoter relative to the exogenous nucleic acid. Furthermore, the exogenous nucleic acid preferably has all appropriate sequences for expression of the nucleic acid. The nucleic acid can include, for example, expression control sequences, such as an enhancer, and necessary information processing sites, such as ribosome binding sites, RNA splice sites, polyadenylation sites, and transcriptional terminator sequences.
[0061]The exogenous nucleic acid can encode beneficial proteins or polypeptides that replace missing or defective proteins required by the cell or subject into which the vector is transferred or can encode a cytotoxic polypeptide that can be directed, e.g., to cancer cells or other cells whose death would be beneficial to the subject. The exogenous nucleic acid can also encode antisense RNAs that can bind to, and thereby inactivate, mRNAs made by the subject that encode harmful proteins. The exogenous nucleic acid can also encode ribozymes that can effect the sequence-specific inhibition of gene expression by the cleavage of mRNAs. In one aspect, antisense polynucleotides can be produced from an exogenous expression cassette in an AAV5 vector construct where the expression cassette contains a sequence that promotes cell-type specific expression (Wirak et al., EMBO 10:289 (1991)). For general methods relating to antisense polynucleotides, see Antisense RNA and DNA, D. A. Melton, Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. (1988).
[0062]Examples of exogenous nucleic acids which can be administered to a cell or subject as part of the present AAVX vector can include, but are not limited to the following: nucleic acids encoding secretory and nonsecretory proteins, nucleic acids encoding therapeutic agents, such as tumor necrosis factors (TNF), such as TNF-α; interferons, such as interferon-α, interferon-β, and interferon-γ, interleukins, such as IL-1, IL-1β, and ILs-2 through -14; GM-CSF; adenosine deaminase; cellular growth factors, such as lymphokines; soluble CD4; Factor VIII; Factor IX; T-cell receptors; LDL receptor; ApoE; ApoC; alpha-1 antitrypsin; ornithine transcarbamylase (OTC); cystic fibrosis transmembrane receptor (CFTR); insulin; Fc receptors for antigen binding domains of antibodies, such as immunoglobulins; anti-HIV decoy tar elements; and antisense sequences which inhibit viral replication, such as antisense sequences which inhibit replication of hepatitis B or hepatitis non-A, non-B virus. The nucleic acid is chosen considering several factors, including the cell to be transfected. Where the target cell is a blood cell, for example, particularly useful nucleic acids to use are those which allow the blood cells to exert a therapeutic effect, such as a gene encoding a clotting factor for use in treatment of hemophilia. Another target cell is the lung airway cell, which can be used to administer nucleic acids, such as those coding for the cystic fibrosis transmembrane receptor, which could provide a gene therapeutic treatment for cystic fibrosis. Other target cells include muscle cells where useful nucleic acids, such as those encoding cytokines and growth factors, can be transduced and the protein the nucleic acid encodes can be expressed and secreted to exert its effects on other cells, tissues and organs, such as the liver. Furthermore, the nucleic acid can encode more than one gene product, limited only, if the nucleic acid is to be packaged in a capsid, by the size of nucleic acid that can be packaged.
[0063]Furthermore, suitable nucleic acids can include those that, when transferred into a primary cell, such as a blood cell, cause the transferred cell to target a site in the body where that cell's presence would be beneficial. For example, blood cells such as TIL cells can be modified, such as by transfer into the cell of a Fab portion of a monoclonal antibody, to recognize a selected antigen. Another example would be to introduce a nucleic acid that would target a therapeutic blood cell to tumor cells. Nucleic acids useful in treating cancer cells include those encoding chemotactic factors which cause an inflammatory response at a specific site, thereby having a therapeutic effect.
[0064]Cells, particularly blood cells, muscle cells, airway epithelial cells, brain cells and endothelial cells having such nucleic acids transferred into them can be useful in a variety of diseases, syndromes and conditions. For example, suitable nucleic acids include nucleic acids encoding soluble CD4, used in the treatment of AIDS and α-antitrypsin, used in the treatment of emphysema caused by α-antitrypsin deficiency. Other diseases, syndromes and conditions in which such cells can be useful include, for example, adenosine deaminase deficiency, sickle cell deficiency, brain disorders such as Alzheimer's disease, thalassemia, hemophilia, diabetes, phenylketonuria, growth disorders and heart diseases, such as those caused by alterations in cholesterol metabolism, and defects of the immune system.
[0065]Other cells in which a gene of interest can be expressed include, but are not limited to, fibroblasts, neurons, retinal cells, kidney cells, lung cells, bone marrow stem cells, hematopoietic stem cells, retinal cells and neurons. The cells in which the gene of interest can be expressed can be dividing cells such as MDCK cells, BHI cells, HeLa cells, 3T3 cells, CV1 cells, COS7 cells, HOS cells and 293 cells. The cells can also be embryonic stem cells of mouse, rhesus, human, bovine or sheep origin, as well as stem cells of neural, hematopoietic, muscle, cardiac, immune or other origin. Non-dividing cells can also be contacted with a particle provided herein to express a gene of interest. Such cells include, but are not limited to hematopoietic stem cells and embryonic stem cells that have been rendered non-dividing.
[0066]As another example, hepatocytes can be transfected with the present vectors having useful nucleic acids to treat liver disease. For example, a nucleic acid encoding OTC can be used to transfect hepatocytes (ex vivo and returned to the liver or in vivo) to treat congenital hyperammonemia, caused by an inherited deficiency in OTC. Another example is to use a nucleic acid encoding LDL to target hepatocytes ex vivo or in vivo to treat inherited LDL receptor deficiency. Such transfected hepatocytes can also be used to treat acquired infectious diseases, such as diseases resulting from a viral infection. For example, transduced hepatocyte precursors can be used to treat viral hepatitis, such as hepatitis B and non-A, non-B hepatitis, for example by transducing the hepatocyte precursor with a nucleic acid encoding an antisense RNA that inhibits viral replication. Another example includes transferring a vector provided herein having a nucleic acid encoding a protein, such as γ-interferon, which can confer resistance to the hepatitis virus.
[0067]For a procedure using transfected hepatocytes or hepatocyte precursors, hepatocyte precursors having a vector provided herein transferred in can be grown in tissue culture, removed from the tissue culture vessel, and introduced to the body, such as by a surgical method. In this example, the tissue would be placed directly into the liver, or into the body cavity in proximity to the liver, as in a transplant or graft. Alternatively, the cells can simply be directly injected into the liver, into the portal circulatory system, or into the spleen, from which the cells can be transported to the liver via the circulatory system. Furthermore, the cells can be attached to a support, such as microcarrier beads, which can then be introduced, such as by injection, into the peritoneal cavity. Once the cells are in the liver, by whatever means, the cells can then express the nucleic acid and/or differentiate into mature hepatocytes which can express the nucleic acid.
[0068]The provided viral particles can be administered to cells, as described herein, with a Multiplicity of Infection (MOI) of 10. The MOI is the ratio of infectious virus particles to the number of cells being infected. Thus, an MOI of 0.1 results in the average inoculation of 1 virus particle for every 10 cells. The general theory behind MOI is to introduce one infectious virus particle to every host cell that is present in the culture. However, more than one virus may infect the same cell which leaves a percentage of cells uninfected. This occurrence can be reduced by using a higher MOI to ensure that every cell is infected. The provided viral particles can therefore be administered to cells, as described herein, with a MOI of 0.01 to 100, such as for example 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 30, 40, 50, 60, 70, 80, 90, 100.
[0069]The AAVX-derived vector can include any normally occurring AAVX nucleic acid sequences. The AAVX-derived vector can also include sequences that are at least 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 98.5%, 99%, 99.5% or 99.9% identical to the AAVX nucleic acids set forth herein. Examples of vector constructs are provided below.
[0070]The present AAVX vector or AAVX particle or recombinant AAVX virion can utilize any unique nucleic acid fragment of the AAVX disclosed herein, including the AAVX nucleic acids set forth in SEQ ID NOS:1-20. A unique fragment consists of a sequence that is not present anywhere else on a genome. A fragment is a subpart of the reference sequence, and thus is identical in sequence to the region of the parent nucleic acid of which it is a fragment. To be unique, the fragment must be of sufficient size to distinguish it from other known sequences, which is most readily determined by comparing any nucleic acid fragment to the nucleotide sequences of nucleic acids in computer databases, such as GenBank. Such comparative searches are standard in the art. Typically, a unique fragment useful as a primer or probe will be at least about 8 or 10, preferable at least 20 or 25 nucleotides in length, depending upon the specific nucleotide content of the sequence. Additionally, fragments can be, for example, at least about 30, 40, 50, 75, 100, 200 or 500 nucleotides in length and can encode polypeptides or be probes. The nucleic acid can be single or double stranded, depending upon the purpose for which it is intended. Where desired, the nucleic acid can be RNA.
[0071]It is understood that as discussed herein the use of the terms “homology” and “identity” mean the same thing as similarity. Thus, for example, if the use of the word homology is used to refer to two non-natural sequences, it is understood that this is not necessarily indicating an evolutionary relationship between these two sequences, but rather is looking at the similarity or relatedness between their nucleic acid sequences. Many of the methods for determining homology between two evolutionarily related molecules are routinely applied to any two or more nucleic acids or proteins for the purpose of measuring sequence similarity regardless of whether they are evolutionarily related.
[0072]In general, it is understood that one way to define any known variants and derivatives or those that might arise, of the disclosed nucleic acids and polypeptides herein, is through defining the variants and derivatives in terms of homology to specific known sequences. In general, variants of nucleic acids and polypeptides herein disclosed typically have at least, about 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 98.5%, 99%, 99.5% or 99.9% homology to the stated sequence or the native sequence. Those of skill in the art readily understand how to determine the homology of two polypeptides or nucleic acids. For example, the homology can be calculated after aligning the two sequences so that the homology is at its highest level.
[0073]Another way of calculating homology can be performed by published algorithms. Optimal alignment of sequences for comparison may be conducted by the local homology algorithm of Smith and Waterman Adv. Appl. Math. 2: 482 (1981), by the homology alignment algorithm of Needleman and Wunsch, J. MoL Biol. 48: 443 (1970), by the search for similarity method of Pearson and Lipman, Proc. Natl. Acad. Sci. U.S.A. 85: 2444 (1988), by computerized implementations of these algorithms (GAP, BESTFIT, FASTA, and TFASTA in the Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, Wis.; the BLAST algorithm of Tatusova and Madden FEMS Microbiol. Lett. 174: 247-250 (1999) available from the National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/blast/b12seq/b12.html), or by inspection.
[0074]The same types of homology can be obtained for nucleic acids by for example the algorithms disclosed in Zuker, M. Science 244:48-52, 1989, Jaeger et al. Proc. Natl. Acad. Sci. USA 86:7706-7710, 1989, Jaeger et al. Methods Enzymol. 183:281-306, 1989, which are herein incorporated by reference for at least material related to nucleic acid alignment. It is understood that any of the methods typically can be used and that in certain instances the results of these various methods may differ, but the skilled artisan understands if identity is found with at least one of these methods, the sequences would be said to have the stated identity.
[0075]For example, as used herein, a sequence recited as having a particular percent homology to another sequence refers to sequences that have the recited homology as calculated by any one or more of the calculation methods described above. For example, a first sequence has 80 percent homology, as defined herein, to a second sequence if the first sequence is calculated to have 80 percent homology to the second sequence using the Zuker calculation method even if the first sequence does not have 80 percent homology to the second sequence as calculated by any of the other calculation methods. As another example, a first sequence has 80 percent homology, as defined herein, to a second sequence if the first sequence is calculated to have 80 percent homology to the second sequence using both the Zuker calculation method and the Pearson and Lipman calculation method even if the first sequence does not have 80 percent homology to the second sequence as calculated by the Smith and Waterman calculation method, the Needleman and Wunsch calculation method, the Jaeger calculation methods, or any of the other calculation methods. As yet another example, a first sequence has 80 percent homology, as defined herein, to a second sequence if the first sequence is calculated to have 80 percent homology to the second sequence using each of calculation methods (although, in practice, the different calculation methods will often result in different calculated homology percentages).
[0076]Further provided herein is an AAVX capsid protein that can combine with other capsid proteins to form an AAVX particle to contain the disclosed vectors. Also provided herein is an AAVX particle, comprising an AAVX capsid protein. The capsid protein can be selected from a group consisting of VP1, VP2 and VP3.
[0077]The AAV-X1 VP1 capsid protein of an AAV-X1 particle can have the amino acid sequence of SEQ ID NO:21. The AAV-X1b VP1 capsid protein of an AAV-X1b particle can have the amino acid sequence of SEQ ID NO:22. The AAV-X5 VP 1 capsid protein of an AAV-X5 particle can have the amino acid sequence of SEQ ID NO:23. The AAV-X19 VP1 capsid protein of an AAV-X19 particle can have the amino acid sequence of SEQ ID NO:24. The AAV-X21 VP1 capsid protein of an AAV-X21 particle can have the amino acid sequence of SEQ ID NO:25. The AAV-X22 VP1 capsid protein of an AAV-X22 particle can have the amino acid sequence of SEQ ID NO:26. The AAV-X23 VP1 capsid protein of an AAV-X23 particle can have the amino acid sequence of SEQ ID NO:27. The AAV-X24 VPI capsid protein of an AAV-X24 particle can have the amino acid sequence of SEQ ID NO:28. The AAV-X25 capsid protein of an AAV-X25 particle can have the amino acid sequence of SEQ ID NO:29. The AAV-X26 capsid protein of an AAV-X26 particle can have the amino acid sequence of SEQ ID NO:30.
[0078]The AAV-X1 VP1 capsid protein of an AAV-X1 particle can be encoded by the nucleic acid sequence of SEQ ID NO:11. The AAV-X1b VP1 capsid protein of an AAV-X1b particle can be encoded by the nucleic acid sequence of SEQ ID NO:12. The AAV-X5 VP1 capsid protein of an AAV-X5 particle can be encoded by the nucleic acid sequence of SEQ ID NO:13. The AAV-X19 VP1 capsid protein of an AAV-X19 particle can be encoded by the nucleic acid sequence of SEQ ID NO:14. The AAV-X21 VP1 capsid protein of an AAV-X21 particle can be encoded by the nucleic acid sequence of SEQ ID NO:15. The AAV-X22 VP1 capsid protein of an AAV-X22 particle can be encoded by the nucleic acid sequence of SEQ ID NO:16. The AAV-X23 VP1 capsid protein of an AAV-X23 particle can be encoded by the nucleic acid sequence of SEQ ID NO:17. The AAV-X24 VP1 capsid protein of an AAV-X24 particle can be encoded by the nucleic acid sequence of SEQ ID NO:18. The AAV-X25 VP1 capsid protein of an AAV-X25 particle can be encoded by the nucleic acid sequence of SEQ ID NO:19. The AAV-X26 VP1 capsid protein of an AAV-X26 particle can be encoded by the nucleic acid sequence of SEQ ID NO:20.
[0079]For example, provided is an AAVX particle, comprising all three AAVX capsid proteins, i.e., VP1, VP2 and VP3. Also provided is an AAVX particle, comprising each AAVX capsid protein individually or in combination. Also provided is an AAVX particle comprising VP1 and VP3 capsid proteins, i.e., lacking any VP2 capsid proteins. Thus, an AAVX particle comprising an AAVX capsid protein comprises at least one AAVX capsid protein (VP1, VP2 or VP3) or a functional fragment thereof. One of skill in the art understands that it is the non-conserved amino acids that are contributing to the properties of AAVX that make it distinct from the other serotypes. Provided therefore is a capsid protein comprising a mutation, deletion or substitution in the conserved regions, including, for example, a substitution with a homologous region from another AAV serotype.,
[0080]An AAVX particle comprising an AAVX capsid protein can be utilized to deliver a nucleic acid vector to a cell, tissue or subject. For example, the herein described AAVX vectors can be encapsidated in an AAVX capsid-derived particle and utilized in a gene delivery method. Furthermore, other viral nucleic acids can be encapsidated in the AAVX particle and utilized in such delivery methods. For example, an AAV1-11, AAAV, or BAAV vector (e.g. AAV1-11, AAAV, BAAV or AAVX ITR and nucleic acid of interest) can be encapsidated in an AAVX particle and administered. Furthermore, an AAVX chimeric capsid incorporating AAV1-11, AAAV, BAAV or AAVX capsid sequences and a different AAVX capsid sequences can be generated, by standard cloning methods, selecting regions from the known sequences of each protein as desired. For example, particularly antigenic regions of the AAVX capsid protein can be replaced with the corresponding region of the AAV2 capsid protein. In addition to chimeric capsids incorporating AAV2 capsid sequences, chimeric capsids incorporating AAV1, 3-8, AAAV, BAAV or AAVX capsid sequences can be generated, by standard cloning methods, selecting regions from the known sequences of each protein as desired. Alternatively a chimeric capsid can be made by the addition of a plasmid that expresses AAV1-11, AAAV, BAAV or AAVX capsid proteins at a ratio with the AAVX capsid expression plasmid that allows only a few capsid proteins to be incorporated into the AAVX particle. Thus, for example, a chimeric particle may be constructed that contains 6 AAV2 capsid proteins and 54 AAVX capsid proteins if the complete capsid contains 60 capsid proteins. Methods for generating chimeric AAVs are known in the art and can be found in Rabinowitz J E, et al. J. Virol. 2004 May; 78(9):4421-32, herein incorporated by reference for these methods. Examples of chimeric capsids would be to combine the VP1, 2, 3 proteins of AAVX and the VP1, 2, 3 proteins of AAV5 such that a new tropism would arise. The capsids can also be modified to alter their specific tropism by genetically altering the capsid to encode a specific ligand to a cell surface receptor.
[0081]Alternatively, the capsid can be chemically modified by conjugating a ligand to a cell surface receptor. By genetically or chemically altering the capsids, the tropism can be modified to direct AAVX to a particular cell or population of cells. The capsids can also be altered immunologically by conjugating the capsid to an antibody that recognizes a specific protein on the target cell or population of cells.
[0082]Provided are three regions in the capsid of AAVX that are on the virus surface and could tolerate substitution. These three regions in AAV-X1, AAV-X1b, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, and AAV-X25 are aa 261-271, aa 450-476, and aa 546-559. These three regions in AAV-X5 are aa 259-268, aa 448-473, and aa 543-554. These three regions in AAV-X26 are aa 260-274, aa 445-477, and aa 550-565. Thus, provided is an AAVX VP1 capsid, comprising amino acid substitutions in aa 261-271, aa 450-476, or aa 546-559 of SEQ ID NOS:21, 22, 24, 25, 26, 27, 28, or 29. Thus, also provided is an AAVX VP1 capsid, comprising amino acid substitutions in aa 259-268, aa 448-473, and aa 543-554 of SEQ ID NO:23. Thus, also provided is an AAVX VP1 capsid, comprising amino acid substitutions in aa 260-274, aa 445-477, and aa 550-565 of SEQ ID NO:30.
[0083]Other regions of the AAVX capsid could also accommodate the substitution of amino acids that would allow for epitope presentation on the surface of the virus. All of these regions would have surface exposure and the ability to support a substitution of sequence to insert the epitope while still allowing for capsid assembly. The substitutions can include non-AAVX epitopes and non-AAVX ligands.
[0084]Because of the symmetry of the AAV particles, a substitution in one subunit of the capsid will appear multiple times on the capsid surface. For example the capsid is made of approximately 50 VP3 proteins, 5 VP1 and 5 VP2. Therefore an epitope incorporated in the VP3 protein could be expressed 55 times on the surface of each particle increasing the likelihood of the epitope forming a stable interaction with its target. In some cases this may be too high of a ligand density for functional binding or this high density of epitope may interfere with capsid formation. The epitope density could be lowered by introducing another plasmid into the packaging system for production of recombinant particles and the ratio between the packaging plasmid with the modified VP3 protein and the wt VP3 protein altered to balance the epitope density on the virus surface. Thus, the ratio between the modified VP3 and the wt VP3 can be 0:50 to 50:0, including, for example, 1:49, 2:48, 3:47, 4:46, 5:45, 6:44, 7:43, 8:42, 9:41, 10:40, 11:39, 12:38, 13:37, 14:36, 15:35, 16:34, 17:33, 18:32, 19:31, 20:30, 21:29, 22:28, 23:27, 24:27, 25:25, 26:24, 27:23, 28:22, 29:21, 30:20, 31:19, 32:18, 33:17, 34:16, 35:15, 36:14, 37:13, 38:12, 39:11, 40:10, 41:9, 42:8, 43:7, 44:6, 45:5, 46:4, 47:3, 48:2, or 49:1.
[0085]Epitopes could be incorporated into the virus capsid for the purpose of 1) altering the tropism of the virus 2) blocking an immune response directed at the virus 3) developing a host immune response to the epitope for the purpose of vaccination.
[0000]Examples of epitopes that could be added to AAVX capsids include but are not limited to
LH receptor binding epitope
RGD integrin binding epitope
CD13 binding epitope NGRAHA SEQ ID NO:35
The Retanefpolyprotein vaccine candidate for HIV-1
single chain antibody fragments directed against tumor cells
Endothelial cell binding epitope SIGYPLP SEQ ID NO:36
serpin receptor ligand, KFNKPFVFLI SEQ ID NO:37
protective B-cell epitope hemagglutinin (HA) 91-108 from influenza HA
NDV B-cell immunodominant epitope (IDE) spanning residues 447 to 455
Major immunogenic epitope for parvovirus B19 (NISLDNPLENPSSLFDLVARIK SEQ ID NO:38) that can elicit protective antibody titers.
[0086]The capsids can also be assembled into empty particles by expression in mammalian, bacterial, fungal or insect cells. For example, AAV2 particles are known to be made from VP3 and VP2 capsid proteins in baculovirus. The same basic protocol can produce an empty AAVX particle comprising AAVX capsid proteins and also full particles. The empty AAVX particles can be used to deliver, for example, antigens, drugs, proteins, or metals to cells or cells in a subject. Antigens can be directly incorporated into the capsid of an empty AAVX particle. An antigen can further be coupled via an antibody-antigen complex to the empty particle. Also disclosed is the coupling of drugs, proteins, or metals on the inside of the empty particles.
[0087]The herein described recombinant AAVX nucleic acid derived vector can be encapsidated in a viral particle. The viral particle can be a parvovirus particle. The parvovirus particle can be a dependovirus particle. The viral particle can be an AAV particle. In particular, the recombinant AAVX nucleic acid derived vector can be encapsidated in an AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, BAAV, AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, or AAV-X26 particle, a particle comprising a portion of any of these capsids, or a chimeric capsid particle as described above, by standard methods using the appropriate capsid proteins in the encapsidation process, as long as the nucleic acid vector fits within the size limitation of the particle utilized. The encapsidation process itself is standard in the art. The AAVX replication machinery, i.e. the rep initiator proteins and other functions required for replication, can be utilized to produce the AAVX genome that can be packaged in an AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, BAAV, AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, or AAV-X26 capsid.
[0088]The recombinant AAVX virion containing a vector can also be produced by recombinant methods utilizing multiple plasmids. In one example, the AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, BAAV, AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, or AAV-X26 rep nucleic acid would be cloned into one plasmid, the AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, BAAV, AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, or AAV-X26 ITR nucleic acid would be cloned into another plasmid and the AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, BAAV, AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, or AAV-X26 capsid nucleic acid would be cloned on another plasmid. These plasmids would then be introduced into cells. The cells that were efficiently transduced by all three plasmids, would exhibit specific integration as well as the ability to produce AAVX recombinant virus. Additionally, two plasmids could be used where the AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, BAAV, AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, or AAV-X26 rep nucleic acid would be cloned into one plasmid and the AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, BAAV, AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, or AAV-X26 ITR and AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, BAAV, AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, or AAV-X26 capsid would be cloned into another plasmid. These plasmids would then be introduced into cells. The cells that were efficiently transduced by both plasmids, would exhibit specific integration as well as the ability to produce AAVX recombinant virus.
[0089]An AAV-X1, AAV-X1b, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, or AAV-X25 capsid can have about 98%, 98.5%, 99%, 99.1%, 99.2%, 99.3%, 99.4%, 99.5%, 99.6%, 99.7%, 99.8%, or 99.9% homology to the polypeptide having the amino acid sequence encoded by nucleotides in SEQ ID NOS:11, 12, 14, 15, 16, 17, 18, or 19, respectively. An AAV-X5 capsid can have about 93%, 94%, 95%, 96%, 97%, 98%, 98.5%, or 99% homology to the polypeptide having the amino acid sequence encoded by nucleotides in SEQ ID NO:13. An AAV-X26 capsid can have about 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 98.5%, or 99% homology to the polypeptide having the amino acid sequence encoded by nucleotides in SEQ ID NO:20.
[0090]An AAV-X1, AAV-X1b, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, or AAV-X25 capsid protein can have about 98%, 98.5%, 99%, 99.1%, 99.2%, 99.3%, 99.4%, 99.5%, 99.6%, 99.7%, 99.8%, or 99.9% homology to the protein having the amino acid sequence encoded by the nucleotides set forth in SEQ ID NOS:21, 22, 24, 25, 26, 27, 28, or 29, respectively. An AAV-X5 capsid protein can have about 93%, 94%, 95%, 96%, 97%, 98%, 98.5%, or 99% homology to the protein having the amino acid sequence encoded by the nucleotides set forth in SEQ ID NO:23. An AAV-X26 capsid protein can have about 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 98.5%, or 99% homology to the protein having the amino acid sequence encoded by the nucleotides set forth in SEQ ID NO:30.
[0091]The percent homology used to identify proteins herein, can be based on a nucleotide-by-nucleotide comparison or more preferable is based on a computerized algorithm as described herein. Variations in the amino acid sequence of the AAVX capsid protein are contemplated herein, as long as the resulting particle comprising an AAVX capsid protein remains antigenically or immunologically distinct from AAV1-11, AAAV, or BAAV capsid, as can be routinely determined by standard methods. Specifically, for example, ELISA and Western blots can be used to determine whether a viral particle is antigenically or immunologically distinct from AAV2 or the other serotypes. Furthermore, the AAVX particle preferably retains tissue tropism distinction from other AAVs, such as that exemplified in the examples herein. An AAVX chimeric particle comprising at least one AAVX coat protein may have a different tissue tropism from that of an AAVX particle consisting only of AAVX coat proteins, but is still distinct from the tropism of an AAV2 particle.
[0092]Provided herein is a recombinant AAVX virion, comprising an AAVX particle containing, i.e., encapsidating, a vector comprising a pair of AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, BAAV, AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, or AAV-X26 inverted terminal repeats. The recombinant vector can further comprise an AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, BAAV, AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, or AAV-X26 Rep-encoding nucleic acid. The vector encapsidated in the particle can further comprise an exogenous nucleic acid inserted between the inverted terminal repeats.
[0093]Further contemplated are chimeric recombinant ITRs that contain a rep binding site and a TRS site recognized by that Rep protein. By “Rep protein” is meant one or more of the Rep proteins, Rep 40, Rep 78, Rep 52, Rep 68. Alternatively, “Rep protein” could be all four of the Rep proteins described herein.
[0094]Examples of the combinations of ITR, Rep protein and Capsids that will produce recombinant virus include but are not limited to:
[0000]AAVX ITR+AAVX Rep+AAVX Cap=virus
AAV5 ITR+AAVX Rep+AAVX Cap=virus
AAV5 ITR+AAVX Rep+AAV1 Cap=virus
AAV5 ITR+AAVX Rep+AAV2 Cap=virus
AAV5 ITR+AAVX Rep+AAV3 Cap=virus
AAV5 ITR+AAVX Rep+AAV4 Cap=virus
AAV5 ITR+AAVX Rep+AAV5 Cap=virus
AAV5 ITR+AAVX Rep+AAV6 Cap=virus
AAV5 ITR+AAVX Rep+AAV7 Cap=virus
AAV5 ITR+AAVX Rep+AAV8 Cap=virus
AAV5 ITR+AAVX Rep+AAV9 Cap=virus
AAV5 ITR+AAVX Rep+AAV10 Cap=virus
AAV5 ITR+AAVX Rep+AAV11 Cap=virus
AAV5 ITR+AAVX Rep+AAAV Cap=virus
AAV5 ITR+AAVX Rep+BAAV Cap=virus
AAVX ITR+AAV5 Rep+AAVX Cap=virus
AAVX ITR+AAV5 Rep+AAV1 Capvirus
[0095]AAVX ITR+AAV5 Rep+AAV2 Cap=virus
AAVX ITR+AAV5 Rep+AAV3 Cap=virus
AAVX ITR+AAV5 Rep+AAV4 Cap=virus
AAVX ITR+AAV5 Rep+AAV5 Cap=virus
AAVX ITR+AAV5 Rep+AAV6 Cap=virus
AAVX ITR+AAV5 Rep+AAV7 Cap=virus
AAVX ITR+AAV5 Rep+AAV8 Cap=virus
AAVX ITR+AAV5 Rep+AAV9 Cap=virus
AAVX ITR+AAV5 Rep+AAV10 Cap=virus
AAVX ITR+AAV5 Rep+AAV11 Cap=virus
AAVX ITR+AAV5 Rep+AAAV Cap=virus
AAVX ITR+AAV5 Rep+BAAV Cap=virus
AAV1 ITR+AAV1 Rep+AAVX Cap=virus
AAV2 ITR+AAV2 Rep+AAVX Cap=virus
AAV3 ITR+AAV3 Rep+AAVX Cap=virus
AAV4 ITR+AAV4 Rep+AAVX Cap=virus
AAV5 ITR+AAV5 Rep+AAVX Cap=virus
AAV6 ITR+AAV6 Rep+AAVX Cap=virus
AAV7 ITR+AAV7 Rep+AAVX Cap=virus
AAV8 ITR+AAV8 Rep+AAVX Cap=virus
AAV9 ITR+AAV9 Rep+AAVX Cap=virus
AAV10 ITR+AAV10 Rep+AAVX Cap=virus
AAV11 ITR+AAV11 Rep+AAVX Cap=virus
AAAV ITR+AAAV Rep+AAVX Cap=virus
BAAV ITR+BAAV Rep+AAVX Cap=virus
[Note that AAVX can be AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, or AAV-X26]
[0096]One of skill in the art would know how to employ standard techniques to obtain the sequences from any of AAV1-11, AAAV, BAAV or AAVX in order to combine them with AAVX sequences. Examples of AAVX sequences that can be utilized in these constructs can be found herein. Examples of AAV1 sequences that can be utilized in these constructs can be found in GenBank under Accession No. AF063497 and these sequences are hereby incorporated in their entireties by this reference. Examples of AAV2 sequences that can be utilized in these constructs can be found in GenBank under Accession No. AF043303 and these sequences are hereby incorporated in their entireties by this reference. Examples of AAV3 sequences that can be utilized in these constructs can be found in GenBank under Accession No. NC—001729 and these sequences are hereby incorporated in their entireties by this reference. Examples of AAV4 sequences that can be utilized in these constructs can be found in GenBank under Accession No. U89790 and these sequences are hereby incorporated in their entireties by this reference. Examples of AAV5 sequences that can be utilized in these constructs can be found in GenBank under Accession No. AF085716 and these sequences are hereby incorporated in their entireties by this reference. Examples of AAV6 sequences that can be utilized in these constructs can be found in GenBank under Accession No. NC—001862 and AF028704 and these sequences are hereby incorporated in their entireties by this reference. Examples of AAV7 sequences that can be utilized in these constructs can be found in GenBank under Accession No. AF513851 and these sequences are hereby incorporated in their entireties by this reference. Examples of AAV8 sequences that can be utilized in these constructs can be found in GenBank under Accession No. AF513852 and these sequences are hereby incorporated in their entireties by this reference. Examples of AAV9 sequences that can be utilized in these constructs can be found in GenBank under Accession No. AY530579 and these sequences are hereby incorporated in their entireties by this reference. Examples of AAV10 sequences that can be utilized in these constructs can be found in GenBank under Accession No. AY631965 and these sequences are hereby incorporated in their entireties by this reference. Examples of AAV11 sequences that can be utilized in these constructs can be found in GenBank under Accession No. AY631966 and these sequences are hereby incorporated in their entireties by this reference. Examples of AAAV sequences that can be utilized in these constructs can be found in GenBank under Accession No. AY186198 and these sequences are hereby incorporated in their entireties by this reference. Examples of BAAV sequences that can be utilized in these constructs can be found in GenBank Accession No. AY388617 and these sequences are hereby incorporated in their entireties by this reference.
[0097]In any of the constructs described herein, inclusion of a promoter is preferred. As used in the constructs herein, unless otherwise specified, Cap (capsid) refers to any of VP1, VP2, VP3, combinations thereof, functional fragments of any of VP1, VP2 or VP3, or chimeric capsids as described herein. The ITRs of the constructs described herein, can be chimeric recombinant ITRs as described elsewhere in the application.
[0098]Conjugates of recombinant or wild-type AAVX virions and nucleic acids or proteins can be used to deliver those molecules to a cell. For example, the purified AAVX can be used as a vehicle for delivering DNA bound to the exterior of the virus. Examples of this are to conjugate the DNA to the virion by a bridge using poly-L-lysine or other charged molecule. Also contemplated are virosomes that contain AAVX structural proteins (AAVX capsid proteins), lipids such as DOTAP, and nucleic acids that are complexed via charge interaction to introduce DNA into cells.
[0099]Also provided herein are AAVX capsid proteins (e.g. VP1, VP2 or VP3 or combinations thereof), or AAVX particles consisting of AAVX capsid proteins, wherein the capsid proteins or particles do not contain AAV nucleic acid, vector or plasmid, and are therefore not infectious. These capsid proteins and “empty particles” can comprise other substances such as biologically active molecules (e.g., small molecules, polypeptides, or non-AAV nucleic acids). The substances can be conjugated to the capsid proteins or comprise a fusion protein with an AAVX capsid polypeptide. Alternatively, the substance can be incorporated within an AAVX empty particle. AAVX capsid proteins and empty particles can be used to deliver the substance to a target cell using the targeting ability of the capsid protein to achieve the desired tissue tropism. In addition, the empty particles can function to protect the substance from depredation or immune response.
[0100]Also provided herein are conjugates that utilize the AAVX capsid or a unique region of the AAVX capsid protein (e.g. VP1, VP2 or VP3 or combinations thereof) to introduce DNA into cells. For example, the AAVX VP1 protein or fragment thereof, can be conjugated to a DNA on a plasmid that is conjugated to a lipid. Cells can be infected using the targeting ability of the VP 1 capsid protein to achieve the desired tissue tropism, specific to the AAVX. AAVX VP2 and VP3 proteins can also be utilized to introduce DNA or other molecules into cells. By further incorporating the Rep protein and the AAV TRS into the DNA-containing conjugate, cells can be transduced and targeted integration can be achieved. For example, if AAVX specific targeted integration is desired, a conjugate composed of the AAVX VP1 capsid, AAVX Rep or a fragment of AAVX Rep, AAVX TRS, the Rep binding site, the exogenous DNA of interest, and a lipid, can be utilized to achieve AAVX specific tropism and AAVX specific targeted integration in the genome.
[0101]Further provided herein are chimeric viruses where AAVX vectors can be encapsidated by herpes simplex virus (HSV) (Heister, T., et al. J. Virol. 2002 July; 76(14):7163-73), incorporated herein for its teaching of HSV/AAV hybrid vectors), baculovirus or other viruses to achieve a desired tropism associated with another virus. For example, the AAVX ITRs could be encapsidated by HSV and cells could be infected. Post-infection, the ITRs of AAVX could be acted on by AAVX Rep provided in the system or in a separate vehicle to rescue AAVX from the genome. Therefore, the cellular tropism of HSV can be combined with AAVX Rep mediated targeted integration. Other viruses that could be utilized to construct chimeric viruses include lentivirus, retrovirus, pseudotyped retroviral vectors and adenoviral vectors.
[0102]Provided herein are isolated nucleic acids of AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, and AAV-X26. For example, provided is an isolated nucleic acid comprising the nucleotide sequence set forth in SEQ ID NO:1 (AAV-X1 partial genome). Also provided is an isolated nucleic acid comprising the nucleotide sequence set forth in SEQ ID NO:2 (AAV-X1b partial genome). Also provided is an isolated nucleic acid comprising the nucleotide sequence set forth in SEQ ID NO:3 (AAV-X5 partial genome). Also provided is an isolated nucleic acid comprising the nucleotide sequence set forth in SEQ ID NO:4 (AAV-X19 partial genome). Also provided is an isolated nucleic acid comprising the nucleotide sequence set forth in SEQ ID NO:5 (AAV-X21 partial genome). Also provided is an isolated nucleic acid comprising the nucleotide sequence set forth in SEQ ID NO:6 (AAV-X22 partial genome). Also provided is an isolated nucleic acid comprising the nucleotide sequence set forth in SEQ ID NO:7 (AAV-X23 partial genome). Also provided is an isolated nucleic acid comprising the nucleotide sequence set forth in SEQ ID NO:8 (AAV-X24 partial genome). Also provided is an isolated nucleic acid comprising the nucleotide sequence set forth in SEQ ID NO:9 (AAV-X25 partial genome). Also provided is an isolated nucleic acid comprising the nucleotide sequence set forth in SEQ ID NO:10 (AAV-X26 partial genome).
[0103]This nucleic acid, or unique portions thereof, can be inserted into vectors, such as plasmids, yeast artificial chromosomes, or other viral vector (particle), if desired, by standard cloning methods. Also provided is an isolated nucleic acid consisting essentially of the nucleotide sequence set forth in SEQ ID NOs:1-10.
[0104]The phrase “consisting essentially of” is used herein to refer to a composition that comprises the essential characteristics of the identified composition. By “essential” is meant the characteristics that contribute to the structure or function of the disclosed molecule. Thus, any substitution, deletion or addition to the provided composition that does not significantly alter the defining characteristics of the composition are considered therein.
[0105]For example, if an amino acid sequence X is disclosed, then a provided polypeptide consisting essentially of the amino acid sequence X includes, for example, conservative amino acid substitutions (as described below) that do not significantly alter the essential characteristics of the polypeptide, e.g., secondary/tertiary structure or function of the protein. The provided polypeptide can further constitute a fusion protein or otherwise have additional N-terminal, C-terminal, or intermediate amino acid sequences, e.g., linkers or tags. “Linker”, as used herein, is an amino acid sequences or insertion that can be used to connect or separate two distinct polypeptides or polypeptide fragments, wherein the linker does not otherwise contribute to the essential function of the composition. A polypeptide provided herein, can have an amino acid linker comprising, for example, the amino acids GLS, ALS, or LLA. A “tag”, as used herein, refers to a distinct amino acid sequence that can be used to detect or purify the provided polypeptide, wherein the tag does not otherwise contribute to the essential function of the composition. The provided polypeptide can further have deleted N-terminal, C-terminal or intermediate amino acids that do not contribute to the essential activity of the polypeptide.
[0106]As another example, if a nucleic acid X is disclosed, then a provided nucleic acid consisting essentially of nucleic acid sequence X, includes, for example, nucleotide substitutions that do not alter the amino acid sequence of the encoded polypeptide, i.e., due to degeneracy. If sequence X comprises introns and exons, then the provided nucleic acid can further be the cDNA sequence that lacks the introns but comprises the exons of sequence X. To the extent that specific genes within a genome are identified herein, it is further understood that the disclosure of a nucleic acid consisting essentially of the genome sequence would include fragments of the genome such as isolated sequences comprising a gene or genes within the genome.
[0107]Other characteristics of nucleic acid or amino acid sequences that are not herein considered essential include, for example, junk DNA between genes or any identifiable sequence unit, e.g., promoters, enhancers, transmembrane domains, poly-adenylation sequences, signal sequences, etc., that when substituted or removed would be presumed by one skilled in the art to not significantly alter the essential characteristics of the disclosed sequence.
[0108]Thus, the nucleotides of SEQ ID NOS:1-10 can have minor modifications and still be contemplated herein. For example, modifications that do not alter the amino acid encoded by any given codon (such as by modification of the third, “wobble,” position in a codon) can readily be made, and such alterations are known in the art. Furthermore, modifications that cause a resulting neutral (conserved) amino acid substitution of a similar amino acid can be made in a coding region of the genome. Additionally, modifications as described herein for the AAVX components, such as the ITRs, the p5 promoter, etc. are contemplated herein. Furthermore, modifications to regions of SEQ ID NOS:1-10, other than in the ITR, TRS, Rep binding site and hairpin, are likely to be tolerated without serious impact on the function of the nucleic acid as a recombinant vector.
[0109]As used herein, the term “isolated” refers to a nucleic acid separated or significantly free from at least some of the other components of the naturally occurring organism, for example, the cell structural components or viral components commonly found associated with nucleic acids in the environment of the virus and/or other nucleic acids. The isolation of the native nucleic acids can be accomplished, for example, by techniques such as cell lysis followed by phenol plus chloroform extraction, followed by ethanol precipitation of the nucleic acids. The nucleic acids provided herein can be isolated from cells according to any of many methods well known in the art.
[0110]As used herein, the term “nucleic acid” refers to single- or multiple-stranded molecules which may be DNA or RNA, or any combination thereof, including modifications to those nucleic acids. The nucleic acid may represent a coding strand or its complement, or any combination thereof. Nucleic acids may be identical in sequence to the sequences which are naturally occurring for any of the genes discussed herein or may include alternative codons which encode the same amino acid as those provided herein, including that which is found in the naturally occurring sequence. These nucleic acids can also be modified from their typical structure. Such modifications include, but are not limited to, methylated nucleic acids, the substitution of a non-bridging oxygen on the phosphate residue with either a sulfur (yielding phosphorothioate deoxynucleotides), selenium (yielding phosphorselenoate deoxynucleotides), or methyl groups (yielding methylphosphonate deoxynucleotides).
[0111]Additionally provided is an isolated nucleic acid that selectively hybridizes with any nucleic acid disclosed herein, including the AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, or AAV-X26 genome (SEQ ID NOS:1-10) and any unique fragment thereof, including the Rep and capsid encoding sequences, promoters and ITRs. Specifically, the nucleic acid can selectively or specifically hybridize to an isolated nucleic acid consisting of the nucleotide sequence set forth in SEQ ID NO:1, 2, 3, 4, 5, 6, 7, 8, 9, or 10. By “selectively hybridizes” as used herein is meant a nucleic acid that hybridizes to one of the disclosed nucleic acids under sufficient stringency conditions without significant hybridization to a nucleic acid encoding an unrelated protein, and particularly, without detectably hybridizing to nucleic acids of other AAVs. Thus, a nucleic acid that selectively hybridizes with a nucleic acid provided herein will not selectively hybridize under stringent conditions with a nucleic acid encoding a different protein or the corresponding protein from a different serotype of the virus, and vice versa. A “specifically hybridizing” nucleic acid is one that hybridizes under stringent conditions to only a nucleic acid found in AAVX. Therefore, nucleic acids for use, for example, as primers and probes to detect or amplify the target nucleic acids are contemplated herein. Nucleic acid fragments that selectively hybridize to any given nucleic acid can be used, e.g., as primers and or probes for further hybridization or for amplification methods (e.g., polymerase chain reaction (PCR), ligase chain reaction (LCR)). Additionally, for example, a primer or probe can be designed that selectively hybridizes with both AAVX and a gene of interest carried within the AAVX vector (i.e., a chimeric nucleic acid).
[0112]Stringency of hybridization is controlled by both temperature and salt concentration of either or both of the hybridization and washing steps. Typically, the stringency of hybridization to achieve selective hybridization involves hybridization in high ionic strength solution (6×SSC or 6×SSPE) at a temperature that is about 12-25° C. below the Tm(the melting temperature at which half of the molecules dissociate from their hybridization partners) followed by washing at a combination of temperature and salt concentration chosen so that the washing temperature is about 5° C. to 20° C. below the Tm. The temperature and salt conditions are readily determined empirically in preliminary experiments in which samples of reference DNA immobilized on filters are hybridized to a labeled nucleic acid of interest and then washed under conditions of different stringencies. Hybridization temperatures are typically higher for DNA-RNA and RNA-RNA hybridizations. The washing temperatures can be used as described above to achieve selective stringency, as is known in the art. (Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y., 1989; Kunkel et al. Methods Enzymol. 1987:154:367, 1987). A preferable stringent hybridization condition for a DNA:DNA hybridization can be at about 68° C. (in aqueous solution) in 6×SSC or 6×SSPE followed by washing at 68° C. Stringency of hybridization and washing, if desired, can be reduced accordingly as the degree of complementarity desired is decreased, and further, depending upon the G-C or A-T richness of any area wherein variability is searched for. Likewise, stringency of hybridization and washing, if desired, can be increased accordingly as homology desired is increased, and further, depending upon the G-C or AT richness of any area wherein high homology is desired, all as known in the art.
[0113]A nucleic acid that selectively hybridizes to any portion of the AAVX genome is contemplated herein. Therefore, a nucleic acid that selectively hybridizes to AAVX can be of longer length than the AAVX genome, it can be about the same length as the AAVX genome or it can be shorter than the AAVX genome. The length of the nucleic acid is limited on the shorter end of the size range only by its specificity for hybridization to AAVX, i.e., once it is too short, typically less than about 5 to 7 nucleotides in length, it will no longer bind specifically to AAVX, but rather will hybridize to numerous background nucleic acids. Additionally contemplated herein is a nucleic acid that has a portion that specifically hybridizes to AAVX and a portion that specifically hybridizes to a gene of interest inserted within AAVX.
[0114]Provided is an isolated nucleic acid comprising an AAVX p5 promoter. Provided is an isolated nucleic acid comprising an AAVX p19 promoter. Provided is an isolated nucleic acid comprising an AAVX p40 promoter. Provided is an isolated nucleic acid comprising an AAVX ITR. Further provided is an isolated nucleic acid encoding an AAVX Rep protein. The AAVX Rep proteins are encoded by open reading frame (ORF) 1 of the AAVX genome. Examples of the AAV Rep proteins include Rep78, Rep68, Rep52 and Rep40. However, it is contemplated that the Rep nucleic acid can encode any one, two, three, or four of the four Rep proteins, in any order.
[0115]Furthermore, minor modifications are contemplated in the nucleic acid, such as silent mutations in the coding sequences, mutations that make neutral or conservative changes in the encoded amino acid sequence, and mutations in regulatory regions that do not disrupt the expression of the gene. Examples of other minor modifications are known in the art. Further modifications can be made in the nucleic acid, such as to disrupt or alter expression of one or more of the Rep proteins in order to, for example, determine the effect of such a disruption; such as to mutate one or more of the Rep proteins to determine the resulting effect, etc.
[0116]Further provided is an isolated nucleic acid encoding an AAVX Capsid protein. Furthermore, provided is a nucleic acid encoding each of the three AAVX capsid proteins, VP1, VP2, and VP3. Thus, provided is an isolated nucleic acid encoding AAVX VP1, a nucleic acid encoding AAVX VP2, and an isolated nucleic acid encoding AAVX VP3. Thus, provided is an isolated nucleic acid encoding the amino acid sequence set forth in SEQ ID NO:21 (AAV-X1 VP1). Thus, provided is an isolated nucleic acid encoding the amino acid sequence set forth in SEQ ID NO:22 (AAV-X1b VP1). Thus, provided is an isolated nucleic acid encoding the amino acid sequence set forth in SEQ ID NO:23 (AAV-X5 VP1). Thus, provided is an isolated nucleic acid encoding the amino acid sequence set forth in SEQ ID NO:24 (AAV-X19 VP1). Thus, provided is an isolated nucleic acid encoding the amino acid sequence set forth in SEQ ID NO:25 (AAV-X21 VP1). Thus, provided is an isolated nucleic acid encoding the amino acid sequence set forth in SEQ ID NO:26 (AAV-X22 VP1). Thus, provided is an isolated nucleic acid encoding the amino acid sequence set forth in SEQ ID NO:27 (AAV-X23 VP1). Thus, provided is an isolated nucleic acid encoding the amino acid sequence set forth in SEQ ID NO:28 (AAV-X24 VP1). Thus, provided is an isolated nucleic acid encoding the amino acid sequence set forth in SEQ ID NO:29 (AAV-X25 VP1). Thus, provided is an isolated nucleic acid encoding the amino acid sequence set forth in SEQ ID NO:30 (AAV-X26 VP1).
[0117]Also specifically provided is an isolated nucleic acid comprising SEQ ID NO:11 (AAV-X1 VP1). Also specifically provided is an isolated nucleic acid comprising SEQ ID NO:12 (AAV-X1b VP1). Also specifically provided is an isolated nucleic acid comprising SEQ ID NO:13 (AAV-X5 VP1). Also specifically provided is an isolated nucleic acid comprising SEQ ID NO:14 (AAV-X19 VP1). Also specifically provided is an isolated nucleic acid comprising SEQ ID NO:15 (AAV-X21 VP1). Also specifically provided is an isolated nucleic acid comprising SEQ ID NO:16 (AAV-X22 VP1). Also specifically provided is an isolated nucleic acid comprising SEQ ID NO:17 (AAV-X23 VP1). Also specifically provided is an isolated nucleic acid comprising SEQ ID NO:18 (AAV-X24 VP1). Also specifically provided is an isolated nucleic acid comprising SEQ ID NO:19 (AAV-X25 VP1). Also specifically provided is an isolated nucleic acid comprising SEQ ID NO:20 (AAV-X26 VP1).
[0118]Minor modifications in the nucleotide sequences encoding the capsid, or coat, proteins are contemplated, as described above for other AAVX nucleic acids. However, in general, a modified nucleic acid encoding a capsid protein will have at least about 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 98.5%, 99%, 99.5% or 99.9% or 100% homology to the capsid nucleic sequences described herein e.g., SEQ ID NOS:11-20, and the capsid polypeptide encoded therein will have overall about 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 98.5%, 99%, 99.5% or 99.9% or 100% homology with the amino acid sequence described herein, e.g., SEQ ID NOS:21-30. Isolated nucleic acids that selectively hybridize with the nucleic acids of SEQ ID NOS:11-20 under the conditions described above are also provided.
[0119]Also provided is a cell containing one or more of the herein described nucleic acids, such as the AAVX genome, AAVX ORF1 and ORF2, each AAVX Rep protein gene, or each AAVX capsid protein gene. Such a cell can be any desired cell and can be selected based upon the use intended. For example, cells can include bacterial cells, yeast cells, insect cells, human HeLa cells and simian Cos cells as well as other human and mammalian cells and cell lines. Primary cultures as well as established cultures and cell lines can be used. Nucleic acids provided herein can be delivered into cells by any selected means, in particular depending upon the target cells. Many delivery means are well-known in the art. For example, electroporation, calcium phosphate precipitation, microinjection, cationic or anionic liposomes, and liposomes in combination with a nuclear localization signal peptide for delivery to the nucleus can be utilized, as is known in the art. Additionally, if the nucleic acids are in a viral particle, the cells can simply be transduced with the virion by standard means known in the art for AAV transduction. Small amounts of the recombinant AAVX virus can be made to infect cells and produce more of itself.
[0120]Provided herein are purified AAVX polypeptides. The term “polypeptide” as used herein refers to a polymer of amino acids and includes full-length proteins and fragments thereof. Thus, “protein,” polypeptide,” and “peptide” are often used interchangeably herein. Substitutions can be selected by known parameters to be neutral (see, e.g., Robinson WE Jr, and Mitchell WM., AIDS 4:S151-S162 (1990)). As will be appreciated by those skilled in the art, also provided herein are those polypeptides having slight variations in amino acid sequences or other properties. Such variations may arise naturally as allelic variations (e.g., due to genetic polymorphism) or may be produced by human intervention (e.g., by mutagenesis of cloned DNA sequences), such as induced point, deletion, insertion and substitution mutants. Minor changes in amino acid sequence are generally preferred, such as conservative amino acid replacements, small internal deletions or insertions, and additions or deletions at the ends of the molecules. Substitutions may be designed based on, for example, the model of Dayhoff, et al. (in Atlas of Protein Sequence and Structure 1978, Nat'l Biomed. Res. Found., Washington, D.C.). These modifications can result in changes in the amino acid sequence, provide silent mutations, modify a restriction site, or provide other specific mutations. The location of any modifications to the polypeptide will often determine its impact on function. Particularly, alterations in regions non-essential to protein function will be tolerated with fewer effects on function. Elsewhere in the application regions of the AAVX proteins are described to provide guidance as to where substitutions, additions or deletions can be made to minimize the likelihood of disturbing the function of the variant.
[0121]Protein variants and derivatives are well understood to those of skill in the art and in can involve amino acid sequence modifications. For example, amino acid sequence modifications typically fall into one or more of three classes: substitutional, insertional or deletional variants. Insertions include amino and/or carboxyl terminal fusions as well as intrasequence insertions of single or multiple amino acid residues. Insertions ordinarily will be smaller insertions than those of amino or carboxyl terminal fusions, for example, on the order of one to four residues. Deletions are characterized by the removal of one or more amino acid residues from the protein sequence. Typically, no more than about from 2 to 6 residues are deleted at any one site within the protein molecule. These variants ordinarily are prepared by site specific mutagenesis of nucleotides in the DNA encoding the protein, thereby producing DNA encoding the variant, and thereafter expressing the DNA in recombinant cell culture. Techniques for making substitution mutations at predetermined sites in DNA having a known sequence are well known, for example M13 primer mutagenesis and PCR mutagenesis. Amino acid substitutions are typically of single residues, but can occur at a number of different locations at once; insertions usually will be on the order of about from 1 to 10 amino acid residues; and deletions will range about from 1 to 30 residues. Deletions or insertions preferably are made in adjacent pairs, i.e. a deletion of 2 residues or insertion of 2 residues. Substitutions, deletions, insertions or any combination thereof may be combined to arrive at a final construct. The mutations must not place the sequence out of reading frame and preferably will not create complementary regions that could produce secondary mRNA structure. Substitutional variants are those in which at least one residue has been removed and a different residue inserted in its place. Such substitutions generally are made in accordance with the following Tables 1 and 2 and are referred to as conservative substitutions.
[0000] |
| Amino Acid Abbreviations |
| Amino Acid | Abbreviations |
| |
| alanine | Ala | A |
| allosoleucine | AIle |
| arginine | Arg | R |
| asparagine | Asn | N |
| aspartic acid | Asp | D |
| cysteine | Cys | C |
| glutamic acid | Glu | E |
| glutamine | Gln | Q |
| glycine | Gly | G |
| histidine | His | H |
| isolelucine | Ile | I |
| leucine | Leu | L |
| lysine | Lys | K |
| phenylalanine | Phe | F |
| proline | Pro | P |
| pyroglutamic acid | pGlu |
| serine | Ser | S |
| threonine | Thr | T |
| tyrosine | Tyr | Y |
| tryptophan | Trp | W |
| valine | Val | V |
| |
[0000] |
| Ala | Ser |
| Arg | Lys; Gln |
| Asn | Gln; His |
| Asp | Glu |
| Cys | Ser |
| Gln | Asn, Lys |
| Glu | Asp |
| Gly | Pro |
| His | Asn; Gln |
| Ile | Leu; Val |
| Leu | Ile; Val |
| Lys | Arg; Gln |
| Met | Leu; Ile |
| Phe | Met; Leu; Tyr |
| Ser | Thr |
| Thr | Ser |
| Trp | Tyr |
| Tyr | Trp; Phe |
| Val | Ile; Leu |
| |
[0122]Substantial changes in function or immunological identity can result from selecting substitutions that are less conservative than those in Table 2, i.e., selecting residues that differ more significantly in their effect on maintaining (a) the structure of the polypeptide backbone in the area of the substitution, for example as a sheet or helical conformation, (b) the charge or hydrophobicity of the molecule at the target site or (c) the bulk of the side chain. The substitutions which in general are expected to produce the greatest changes in the protein properties will be those in which (a) a hydrophilic residue, e.g. seryl or threonyl, is substituted for (or by) a hydrophobic residue, e.g. leucyl, isoleucyl, phenylalanyl, valyl or alanyl; (b) a cysteine or proline is substituted for (or by) any other residue; (c) a residue having an electropositive side chain, e.g., lysyl, arginyl, or histidyl, is substituted for (or by) an electronegative residue, e.g., glutamyl or aspartyl; or (d) a residue having a bulky side chain, e.g., phenylalanine, is substituted for (or by) one not having a side chain, e.g., glycine, in this case, (e) by increasing the number of sites for sulfation and/or glycosylation.
[0123]For example, the replacement of one amino acid residue with another that is biologically and/or chemically similar is known to those skilled in the art as a conservative substitution. For example, a conservative substitution would be replacing one hydrophobic residue for another, or one polar residue for another. The substitutions include combinations such as, for example, Gly, Ala; Val, Ile, Leu; Asp, Glu; Asn, Gln; Ser, Thr; Lys, Arg; and Phe, Tyr. Such conservatively substituted variations of each explicitly disclosed sequence are included within the mosaic polypeptides provided herein.
[0124]Generally, a conservative substitution is a substitution of an amino acid residue for another amino acid residue having similar biochemical properties. Typically, conservative substitutions have little to no impact on the biological activity of a resulting polypeptide. In a particular example, a conservative substitution is an amino acid substitution in a peptide that does not substantially affect the biological function of the peptide. A peptide can include one or more amino acid substitutions, for example 2-10 conservative substitutions, 2-5 conservative substitutions, 4-9 conservative substitutions, such as 2, 5 or 10 conservative substitutions.
[0125]For example, a conservative substitution in an AAVX VP1 peptide (such as a peptides encoded by SEQ ID NOS:21-30) does not substantially affect the ability of VP1 peptide to confer the unique tropism of the AAVX particle. A polypeptide can be produced to contain one or more conservative substitutions by manipulating the nucleotide sequence that encodes that polypeptide using, for example, standard procedures such as site-directed mutagenesis or PCR. Alternatively, a polypeptide can be produced to contain one or more conservative substitutions by using standard peptide synthesis methods. An alanine scan can be used to identify which amino acid residues in a protein can tolerate an amino acid substitution. In one example, the biological activity of the protein is not decreased by more than 25%, for example not more than 20%, for example not more than 10%, when an alanine, or other conservative amino acid (such as those listed below), is substituted for one or more native amino acids.
[0126]Examples of amino acids which can be substituted for an original amino acid in a protein and which are regarded as conservative substitutions include, but are not limited to: Ser for Ala; Lys for Arg; Gln or H is for Asn; Glu for Asp; Ser for Cys; Asn for Gln; Asp for Glu; Pro for Gly; Asn or Gln for H is; Leu or Val for Ile; Ile or Val for Leu; Arg or Gln for Lys; Leu or Ile for Met; Met, Leu or Tyr for Phe; Thr for Ser; Ser for Thr; Tyr for Trp; Trp or Phe for Tyr; and Ile or Leu for Val.
[0127]Further information about conservative substitutions can be found in, among other locations in, Ben-Bassat et al., (J. Bacteriol. 169:751-7, 1987), O'Regan et al., (Gene 77:237-51, 1989), Sahin-Toth et al., (Protein Sci. 3:240-7, 1994), Hochuli et al., (Bio/Technology 6:1321-5, 1988) and in standard textbooks of genetics and molecular biology.
[0128]Substitutional or deletional mutagenesis can be employed to insert sites for N-glycosylation (Asn-X-Thr/Ser) or O-glycosylation (Ser or Thr). Deletions of cysteine or other labile residues also may be desirable. Deletions or substitutions of potential proteolysis sites, e.g. Arg, is accomplished for example by deleting one of the basic residues or substituting one by glutaminyl or histidyl residues.
[0129]Certain post-translational derivatizations are the result of the action of recombinant host cells on the expressed polypeptide. Glutaminyl and asparaginyl residues are frequently post-translationally deamidated to the corresponding glutamyl and asparyl residues. Alternatively, these residues are deamidated under mildly acidic conditions. Other post-translational modifications include hydroxylation of proline and lysine, phosphorylation of hydroxyl groups of seryl or threonyl residues, methylation of the o-amino groups of lysine, arginine, and histidine side chains (T. E. Creighton, Proteins: Structure and Molecular Properties, W.H. Freeman & Co., San Francisco pp 79-86 [1983]), acetylation of the N-terminal amine and, in some instances, amidation of the C-terminal carboxyl.
[0130]It is understood that there are numerous amino acid and peptide analogs which can be incorporated into the disclosed compositions. For example, there are numerous D amino acids or amino acids which have a different functional substituent then the amino acids shown in Table 1 and Table 2. The opposite stereo isomers of naturally occurring peptides are disclosed, as well as the stereo isomers of peptide analogs. These amino acids can readily be incorporated into polypeptide chains by charging tRNA molecules with the amino acid of choice and engineering genetic constructs that utilize, for example, amber codons, to insert the analog amino acid into a peptide chain in a site specific way (Thorson et al., Methods in Molec. Biol. 77:43-73 (1991), Zoller, Current Opinion in Biotechnology, 3:348-354 (1992); Ibba, Biotechnology & Genetic Engineering Reviews 13:197-216 (1995), Cahill et al., TIBS, 14(10):400-403 (1989); Benner, TIB Tech, 12:158-163 (1994); Ibba and Hennecke, Bio/technology, 12:678-682 (1994) all of which are herein incorporated by reference at least for material related to amino acid analogs).
[0131]Molecules can be produced that resemble peptides, but which are not connected via a natural peptide linkage. For example, linkages for amino acids or amino acid analogs can include CH2NH—, —CH2S—, —CH2—CH2—CH═CH—(cis and trans), —COCH2—CH(OH)CH2—, and —CHH2SO (These and others can be found in Spatola, A. F. in Chemistry and Biochemistry of Amino Acids, Peptides, and Proteins, B. Weinstein, eds., Marcel Dekker, New York, p. 267 (1983); Spatola, A. F., Vega Data (March 1983), Vol. 1, Issue 3, Peptide Backbone Modifications (general review); Morley, Trends Pharm Sci (1980) pp. 463-468; Hudson, D. et al., Int J Pept Prot Res 14:177-185 (1979) (—CH2NH—, CH2CH2—); Spatola et al. Life Sci 38:1243-1249 (1986) (—CH H2—S); Hann J. Chem. Soc Perkin Trans. I 307-314 (1982) (—CH—CH—, cis and trans); Almquist et al. J. Med. Chem. 23:1392-1398 (1980) (—COCH2—); Jennings-White et al. Tetrahedron Lett 23:2533 (1982) (—COCH2—); Szelke et al. European Appin, EP 45665 CA (1982): 97:39405 (1982) (—CH(OH)CH2—); Holladay et al. Tetrahedron. Lett 24:4401-4404 (1983) (—C(OH)CH2—); and Hruby Life Sci 31:189-199 (1982) (—CH2—S—); each of which is incorporated herein by reference. A particularly preferred non-peptide linkage is —CH2NH—. It is understood that peptide analogs can have more than one atom between the bond atoms, such as b-alanine, g-aminobutyric acid, and the like.
[0132]Amino acid analogs and analogs and peptide analogs often have enhanced or desirable properties, such as, more economical production, greater chemical stability, enhanced pharmacological properties (half-life, absorption, potency, efficacy, etc.), altered specificity (e.g., a broad-spectrum of biological activities), reduced antigenicity, and others.
[0133]D-amino acids can be used to generate more stable peptides, because D amino acids are not recognized by peptidases and such. Systematic substitution of one or more amino acids of a consensus sequence with a D-amino acid of the same type (e.g., D-lysine in place of L-lysine) can be used to generate more stable peptides. Cysteine residues can be used to cyclize or attach two or more peptides together. This can be beneficial to constrain peptides into particular conformations. (Rizo and Gierasch Ann. Rev. Biochem. 61:387 (1992), incorporated herein by reference).
[0134]A polypeptide provided herein can be readily obtained by any of several means. For example, the polypeptide of interest can be synthesized chemically by standard methods. Additionally, the coding regions of the genes can be recombinantly expressed and the resulting polypeptide isolated by standard methods. Furthermore, an antibody specific for the resulting polypeptide can be raised by standard methods (see, e.g., Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y., 1988), and the protein can be isolated from a cell expressing the nucleic acid encoding the polypeptide by selective hybridization with the antibody. This protein can be purified to the extent desired by standard methods of protein purification (see, e.g., Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y., 1989).
[0135]An antigenic or immunoreactive fragment of the provided compositions and methods is typically an amino acid sequence of at least about 5 consecutive amino acids, and it can be derived from the AAVX polypeptide amino acid sequence. An antigenic AAVX fragment is any fragment unique to the AAVX protein, as described herein, against which an AAVX-specific antibody can be raised, by standard methods. Thus, the resulting antibody-antigen reaction should be specific for AAVX.
[0136]By “unique fragment thereof” is meant any smaller polypeptide fragment encoded by an AAVX rep gene that is of sufficient length to be found only in the Rep polypeptide. Substitutions and modifications of the amino acid sequence can be made as described herein and, further, can include protein processing modifications, such as glycosylation, to the polypeptide. Typically, to be unique, a polypeptide fragment provided herein will be at least about 5 amino acids in length; however, unique fragments can be 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100 or more amino acids in length. A unique polypeptide will typically comprise such a unique fragment; however, a unique polypeptide can also be determined by its overall homology. A unique polypeptide can be 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100 or more amino acids in length. Uniqueness of a polypeptide fragment can readily be determined by standard methods such as searches of computer databases of known peptide or nucleic acid sequences or by hybridization studies to the nucleic acid encoding the protein or to the protein itself, as known in the art. The uniqueness of a polypeptide fragment can also be determined immunologically as well as functionally. Uniqueness can be simply determined in an amino acid-by-amino acid comparison of the polypeptides.
[0137]Provided is an isolated AAVX Rep protein. An AAVX Rep polypeptide is encoded by ORF1 of AAVX. Also provided is each individual AAVX Rep protein. Provided is an isolated polypeptide, comprising AAVX Rep 52, or a unique fragment thereof. Provided is an isolated polypeptide, comprising AAV Rep 78, or a unique fragment thereof.
[0138]Further provided is an isolated AAVX Capsid protein or a unique fragment thereof. AAVX capsid protein is encoded by ORF 2 of AAVX. Further provided are the individual AAVX capsid proteins, VP1, VP2 and VP3 or unique fragments thereof. Thus, provided is an isolated polypeptide having the amino acid sequence set forth in SEQ ID NO:21 (AAV-X1 VP1). Also provided is an isolated polypeptide having the amino acid sequence set forth in SEQ ID NO:22 (AAV-X1b VP1). Also provided is an isolated polypeptide having the amino acid sequence set forth in SEQ ID NO:23 (AAV-X5 VP1). Also provided is an isolated polypeptide having the amino acid sequence set forth in SEQ ID NO:24 (AAV-X19 VP1). Also provided is an isolated polypeptide having the amino acid sequence set forth in SEQ ID NO:25 (AAV-X21 VP1). Also provided is an isolated polypeptide having the amino acid sequence set forth in SEQ ID NO:26 (AAV-X22 VP1). Also provided is an isolated polypeptide having the amino acid sequence set forth in SEQ ID NO:27 (AAV-X23 VP1). Also provided is an isolated polypeptide having the amino acid sequence set forth in SEQ ID NO:28 (AAV-X24 VP1). Also provided is an isolated polypeptide having the amino acid sequence set forth in SEQ ID NO:29 (AAV-X25 VP1). Also provided is an isolated polypeptide having the amino acid sequence set forth in SEQ ID NO:30 (AAV-X26 VP1). Further provided is an isolated polypeptide consisting essentially of the amino acid sequence set forth in SEQ ID NO:21, 22, 23, 24, 25, 26, 27, 28, 29, or 30.
[0139]By “unique fragment thereof” is meant any smaller polypeptide fragment encoded by any AAVX capsid gene that is of sufficient length to be found only in the AAVX capsid protein. Substitutions and modifications of the amino acid sequence can be made as described above and, further, can include protein processing modifications, such as glycosylation, to the polypeptide. However, an AAVX Capsid polypeptide including all three coat proteins will have greater than about 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 98.5%, 99%, 99.5% or 99.9% overall homology to the polypeptide encoded by the nucleotides set forth in SEQ ID NOS:21-30.
[0140]Also provided herein are isolated AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, and AAV-X26 viruses. In one aspect, the isolated viruses can be used to produce antibodies specific for each AAVX. Thus, provided is an isolated antibody that specifically binds an AAVX-specific protein. The isolated viruses can be used to detect antibodies specific for each AAVX.
[0141]Thus, provided is an isolated antibody that specifically binds an AAVX Rep protein, or a unique epitope thereof. Thus, also provided is an isolated antibody that specifically bind AAVX Rep 52 or AAVX Rep 78, or a unique fragment thereof. Additionally provided is an isolated antibody that specifically binds any of the AAVX capsid proteins (VP1, VP2 or VP3), a unique epitope thereof, or the polypeptide comprising all three AAVX coat proteins. Also provided is an isolated antibody that specifically binds the AAVX capsid protein having the amino acid sequence set forth in SEQ ID NO:21 (AAV-X1 VP1), or that specifically binds a unique fragment thereof. Also provided is an isolated antibody that specifically binds the AAVX capsid protein having the amino acid sequence set forth in SEQ ID NO:22 (AAV-X1b VP1), or that specifically binds a unique fragment thereof. Also provided is an isolated antibody that specifically binds the AAVX capsid protein having the amino acid sequence set forth in SEQ ID NO:23 (AAV-X5 VP1), or that specifically binds a unique fragment thereof. Also provided is an isolated antibody that specifically binds the AAVX capsid protein having the amino acid sequence set forth in SEQ ID NO:24 (AAV-X19 VP1), or that specifically binds a unique fragment thereof. Also provided is an isolated antibody that specifically binds the AAVX capsid protein having the amino acid sequence set forth in SEQ ID NO:25 (AAV-X21 VP1), or that specifically binds a unique fragment thereof. Also provided is an isolated antibody that specifically binds the AAVX capsid protein having the amino acid sequence set forth in SEQ ID NO:26 (AAV-X22 VP1), or that specifically binds a unique fragment thereof. Also provided is an isolated antibody that specifically binds the AAVX capsid protein having the amino acid sequence set forth in SEQ ID NO:27 (AAV-X23 VP1), or that specifically binds a unique fragment thereof. Also provided is an isolated antibody that specifically binds the AAVX capsid protein having the amino acid sequence set forth in SEQ ID NO:28 (AAV-X24 VP1), or that specifically binds a unique fragment thereof. Also provided is an isolated antibody that specifically binds the AAVX capsid protein having the amino acid sequence set forth in SEQ ID NO:29 (AAV-X25 VP1), or that specifically binds a unique fragment thereof. Also provided is an isolated antibody that specifically binds the AAVX capsid protein having the amino acid sequence set forth in SEQ ID NO:30 (AAV-X26 VP1), or that specifically binds a unique fragment thereof. Again, any given antibody can recognize and bind one of a number of possible epitopes present in the polypeptide; thus only a unique portion of a polypeptide (having the epitope) needs to be present in an assay to determine if the antibody specifically binds the polypeptide.
[0142]The antibody can be a component of a composition that comprises an antibody that specifically binds the AAVX protein. The composition can further comprise, e.g., serum, serum-free medium, or a pharmaceutically acceptable carrier such as physiological saline, etc.
[0143]By “an antibody that specifically binds” an AAVX polypeptide or protein is meant an antibody that selectively binds to an epitope on any portion of the AAVX peptide such that the antibody binds specifically to the corresponding AAVX polypeptide without significant background. Specific binding by an antibody further means that the antibody can be used to selectively remove the target polypeptide from a sample comprising the polypeptide or and can readily be determined by radioimmunoassay (RIA), bioassay, or enzyme-linked immunosorbant (ELISA) technology. An ELISA method effective for the detection of the specific antibody-antigen binding can, for example, be as follows: (1) bind the antibody to a substrate; (2) contact the bound antibody with a sample containing the antigen; (3) contact the above with a secondary antibody bound to a detectable moiety (e.g., horseradish peroxidase enzyme or alkaline phosphatase enzyme); (4) contact the above with the substrate for the enzyme; (5) contact the above with a color reagent; (6) observe the color change.
[0144]An antibody can include antibody fragments such as Fab fragments which retain the binding activity. Antibodies can be made as described in, e.g., Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. (1988). Briefly, purified antigen can be injected into an animal in an amount and in intervals sufficient to elicit an immune response. Antibodies can either be purified directly, or spleen cells can be obtained from the animal. The cells are then fused with an immortal cell line and screened for antibody secretion. Individual hybridomas are then propagated as individual clones serving as a source for a particular monoclonal antibody.
[0145]Additionally provided is a method of screening a cell for infectivity by AAVX, comprising contacting the cell with AAVX and detecting the presence of AAVX in the cells. AAVX particles can be detected using any standard physical or biochemical methods. For example, physical methods that can be used for this detection include DNA based methods such as 1) polymerase chain reaction (PCR) for viral DNA or RNA or 2) direct hybridization with labeled probes, and immunological methods such as by 3) antibody directed against the viral structural or non-structural proteins. Catalytic methods of viral detection include, but are not limited to, detection of site and strand specific DNA nicking activity of Rep proteins or replication of an AAV origin-containing substrate. Reporter genes can also be utilized to detect cells that transduce AAVX. For example, β-gal, green fluorescent protein or luciferase can be inserted into a recombinant AAVX. The cell can then be contacted with the recombinant AAVX, either in vitro or in vivo and a colorimetric assay could detect a color change in the cells that would indicate transduction of AAVX in the cell. Additional detection methods are outlined in Fields, Virology, Raven Press, New York, N.Y. 1996.
[0146]Provided is a method of screening a cell for infectivity by AAVX, wherein the presence of AAVX in the cells is determined by nucleic acid hybridization methods, a nucleic acid probe for such detection can comprise, for example, a unique fragment of any of the AAVX nucleic acids provided herein. The uniqueness of any nucleic acid probe can readily be determined as described herein. Additionally, the presence of AAVX in cells can be determined by fluorescence, antibodies to gene products, focus forming assays, plaque lifts, Western blots and chromogenic assays. The nucleic acid can be, for example, the nucleic acid whose nucleotide sequence is set forth in SEQ ID NOS:1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, or 20 or a unique fragment thereof.
[0147]Provided is a method of determining the suitability of an AAVX vector for administration to a subject comprising contacting an antibody-containing sample from the subject with an antigenic fragment of an isolated AAVX Rep or Capsid protein, and detecting an antibody-antigen reaction in the sample, the presence of a neutralizing reaction indicating the AAVX vector to be unsuitable for use in the subject. Further provided is a method of determining the presence in a subject of an AAVX-specific antibody comprising contacting an antibody-containing sample from the subject with an antigenic fragment of an isolated AAVX Rep or Capsid protein and detecting an antibody-antigen reaction in the sample, the presence of a reaction indicating the presence of an AAVX-specific antibody in the subject. The present methods of determining the suitability of an AAVX vector for administration to a subject or the presence of an AAVX-specific antibody in a subject can comprise contacting an antibody-containing sample from the subject with a unique antigenic or immunogenic fragment of an AAVX Rep protein (e.g. Rep 52, Rep 78) and detecting an antibody-antigen reaction in the sample, the presence of a reaction indicating the presence of an AAVX-specific antibody and therefore the AAVX vector to be unsuitable for use in the subject. The AAVX Rep proteins are provided herein, and their antigenic fragments are routinely determined. The AAVX capsid protein can be used to select an antigenic or immunogenic fragment, for example from the amino acid sequence set forth in SEQ ID NO:21 (AAV-X1 VP1), the amino acid sequence set forth in SEQ ID NO:22 (AAV-X1b VP1), the amino acid sequence set forth in SEQ ID NO:23 (AAV-X5 VP1), the amino acid sequence set forth in SEQ ID NO:24 (AAV-X19 VP1), the amino acid sequence set forth in SEQ ID NO:25 (AAV-X21 VP1), the amino acid sequence set forth in SEQ ID NO:26 (AAV-X22 VP1), the amino acid sequence set forth in SEQ ID NO:27 (AAV-X23 VP1, the amino acid sequence set forth in SEQ ID NO:28 (AAV-X24 VP1), the amino acid sequence set forth in SEQ ID NO:29 (AAV-X25 VP1), or the amino acid sequence set forth in SEQ ID NO:30 (AAV-X26 VP1)
[0148]Alternatively, or additionally, an antigenic or immunogenic fragment of an isolated AAVX Rep protein can be utilized in this determination method. Any given antibody can recognize and bind one of a number of possible epitopes present in the polypeptide; thus only a unique portion of a polypeptide (having the epitope) may need to be present in an assay to determine if the antibody specifically binds the polypeptide.
[0149]The AAVX polypeptide fragments can be analyzed to determine their antigenicity, immunogenicity and/or specificity. Briefly, various concentrations of a putative immunogenically specific fragment are prepared and administered to a subject and the immunological response (e.g., the production of antibodies or cell mediated immunity) of an animal to each concentration is determined. The amounts of antigen administered depend on the subject, e.g. a human, rabbit or a guinea pig, the condition of the subject, the size of the subject, etc. Thereafter an animal so inoculated with the antigen can be exposed to the AAVX viral particle or AAVX protein to test the immunoreactivity or the antigenicity of the specific immunogenic fragment. The specificity of a putative antigenic or immunogenic fragment can be ascertained by testing sera, other fluids or lymphocytes from the inoculated animal for cross reactivity with other closely related viruses, such as AAV1-11, AAAV, or BAAV.
[0150]By the “suitability of an AAVX vector for administration to a subject” is meant a determination of whether the AAVX vector will elicit a neutralizing immune response upon administration to a particular subject. A vector that does not elicit a significant immune response is a potentially suitable vector, whereas a vector that elicits a significant, neutralizing immune response (e.g. at least 90%) is thus likely to be unsuitable for use in that subject. Significance of any detectable immune response is a standard parameter understood by the skilled artisan in the field. For example, one can incubate the subject's serum with the virus, then determine whether that virus retains its ability to transduce cells in culture. If such virus cannot transduce cells in culture, the vector likely has elicited a significant immune response.
[0151]Alternatively, or additionally, one skilled in the art could determine whether or not AAVX administration would be suitable for a particular cell type of a subject. For example, the artisan could culture muscle cells in vitro and transduce the cells with AAVX in the presence or absence of the subject's serum. If there is a reduction in transduction efficiency, this could indicate the presence of a neutralizing antibody or other factors that may inhibit transduction. Normally, greater than 90% inhibition would have to be observed in order to rule out the use of AAVX as a vector. However, this limitation could be overcome by treating the subject with an immunosuppressant that could block the factors inhibiting transduction.
[0152]As will be recognized by those skilled in the art, numerous types of immunoassays are available for use in the present methods to detect binding between an antibody and an AAVX polypeptide as provided herein. For instance, direct and indirect binding assays, competitive assays, sandwich assays, and the like, as are generally described in, e.g., U.S. Pat. Nos. 4,642,285; 4,376,110; 4,016,043; 3,879,262; 3,852,157; 3,850,752; 3,839,153; 3,791,932; and Harlow and Lane, Antibodies, A Laboratory Manual, Cold Spring Harbor Publications, N.Y. (1988). For example, enzyme immunoassays such as immunofluorescence assays (IFA), enzyme linked immunosorbent assays (ELISA) and immunoblotting can be readily adapted to accomplish the detection of the antibody. An ELISA method effective for the detection of the antibody bound to the antigen can, for example, be as follows: (1) bind the antigen to a substrate; (2) contact the bound antigen with a fluid or tissue sample containing the antibody; (3) contact the above with a secondary antibody specific for the antigen and bound to a detectable moiety (e.g., horseradish peroxidase enzyme or alkaline phosphatase enzyme); (4) contact the above with the substrate for the enzyme; (5) contact the above with a color reagent; (6) observe color change.
[0153]The antibody-containing sample of this method can comprise any biological sample which would contain the antibody or a cell containing the antibody, such as blood, plasma, serum, bone marrow, saliva and urine.
[0154]Also provided is a method of producing the AAVX virus by transducing a cell with the nucleic acid encoding the virus.
[0155]The present method further provides a method of delivering an exogenous nucleic acid to a cell comprising administering to the cell an AAVX particle containing a vector comprising the nucleic acid inserted between a pair of AAV inverted terminal repeats, thereby delivering the nucleic acid to the cell.
[0156]The AAV ITRs in the vector for the herein described delivery methods can be AAVX ITRs. Furthermore, the AAV ITRs in the vector for the herein described nucleic acid delivery methods can also comprise AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, or BAAV inverted terminal repeats.
[0157]Also provided is a method of delivering an exogenous nucleic acid to a subject comprising administering to a cell of or from the subject an AAVX particle containing a vector comprising the nucleic acid inserted between a pair of AAV inverted terminal repeats, and returning the cell to the subject, thereby delivering the nucleic acid to the subject. The AAV ITRs can be any AAV ITRs, including AAVX ITRs, AAV5 ITRs and AAV2 ITRs. For example, in an ex vivo administration, cells are isolated from a subject by standard means according to the cell type and placed in appropriate culture medium, again according to cell type (see, e.g., ATCC catalog). Viral particles are then contacted with the cells as described above, and the virus is allowed to transduce the cells. Cells can then be transplanted back into the subject's body, again by means standard for the cell type and tissue (e.g., in general, U.S. Pat. No. 5,399,346; for neural cells, Dunnett, S. B. and Björklund, A., eds., Transplantation: Neural Transplantation—A Practical Approach, Oxford University Press, Oxford (1992)). If desired, prior to transplantation, the cells can be studied for degree of transduction by the virus, by known detection means and as described herein. Cells for ex vivo transduction followed by transplantation into a subject can be selected from those listed above, or can be any other selected cell. Preferably, a selected cell type is examined for its capability to be transfected by AAVX. Preferably, the selected cell will be a cell readily transduced with AAVX particles; however, depending upon the application, even cells with relatively low transduction efficiencies can be useful, particularly if the cell is from a tissue or organ in which even production of a small amount of the protein or antisense RNA encoded by the vector will be beneficial to the subject.
[0158]Further provided is a method of delivering an exogenous nucleic acid to a cell in a subject comprising administering to the subject an AAVX particle containing a vector comprising the nucleic acid inserted between a pair of AAV inverted terminal repeats, thereby delivering the nucleic acid to a cell in the subject. Administration can be an ex vivo administration directly to a cell removed from a subject, such as any of the cells listed above, followed by replacement of the cell back into the subject, or administration can be in vivo administration to a cell in the subject. For ex vivo administration, cells are isolated from a subject by standard means according to the cell type and placed in appropriate culture medium, again according to cell type (see, e.g., ATCC catalog). Viral particles are then contacted with the cells as described above, and the virus is allowed to transfect the cells. Cells can then be transplanted back into the subject's body, again by means standard for the cell type and tissue (e.g., for neural cells, Dunnett, S. B. and Björklund, A., eds., Transplantation: Neural Transplantation—A Practical Approach, Oxford University Press, Oxford (1992)). If desired, prior to transplantation, the cells can be studied for degree of transfection by the virus, by known detection means and as described herein.
[0159]Further provided is a method of delivering a nucleic acid to a cell in a subject having neutralizing antibodies to AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, or BAAV comprising administering to the subject an AAVX particle containing a vector comprising the nucleic acid, thereby delivering the nucleic acid to a cell in the subject. A subject that has neutralizing antibodies to AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, or BAAV can readily be determined by any of several known means, such as contacting AAV 1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, or BAAV protein(s) with an antibody-containing sample, such as blood, from a subject and detecting an antigen-antibody reaction in the sample. Delivery of the AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, or BAAV particle can be by either ex vivo or in vivo administration as herein described. Thus, a subject who might have an adverse immunogenic reaction to a vector administered in an AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, or BAAV viral particle can have a desired nucleic acid delivered using an AAVX particle. This delivery system can be particularly useful for subjects who have received therapy utilizing AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, or BAAV particles in the past and have developed antibodies to AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, or BAAV. An AAVX regimen can now be substituted to deliver the desired nucleic acid.
[0160]In any of the methods of delivering exogenous nucleic acids to a cell or subject described herein, the AAVX-conjugated nucleic acid or AAVX particle-conjugated nucleic acids described herein can be used.
[0161]In vivo administration to a human subject or an animal model can be by any of many standard means for administering viruses, depending upon the target organ, tissue or cell. Virus particles can be administered orally, parenterally (e.g., intravenously), by intramuscular injection, intrarectally, by direct tissue or organ injection, by intraperitoneal injection, topically, transdermally, via aerosol delivery, via the mucosa or the like. Viral nucleic acids (non-encapsidated) can also be administered, e.g., as a complex with cationic liposomes, or encapsulated in anionic liposomes. The present compositions can include various amounts of the selected viral particle or non-encapsidated viral nucleic acid in combination with a pharmaceutically acceptable carrier and, in addition, if desired, may include other medicinal agents, pharmaceutical agents, carriers, adjuvants, diluents, etc. Parental administration, if used, is generally characterized by injection. Injectables can be prepared in conventional forms, either as liquid solutions or suspensions, solid forms suitable for solution or suspension in liquid prior to injection, or as emulsions. Dosages will depend upon the mode of administration, the disease or condition to be treated, and the individual subject's condition, but will be that dosage typical for and used in administration of other AAV vectors, such as AAV2 vectors. Often a single dose can be sufficient; however, the dose can be repeated if desirable. Administration methods for gene delivery to the cochlea are routine and are described in Jero, J. et al. (Gene Ther. 2001 Mar. 20; 12(5):539-48) and Staecker H, et al. (Acta Otolaryngol. 2001 January; 121(2):157-63), both references herein incorporated by reference for these methods.
[0162]Administration methods can be used to treat brain disorders such as Parkinson's disease, Alzheimer's disease, and demyelination disease. Other diseases that can be treated by these methods include metabolic disorders such as musculoskeletal diseases, cardiovascular disease, cancer, and autoimmune disorders.
[0163]Administration of this recombinant AAVX virion to the cell can be accomplished by any means, including simply contacting the particle, optionally contained in a desired liquid such as tissue culture medium, or a buffered saline solution, with the cells. The virion can be allowed to remain in contact with the cells for any desired length of time, and typically the virion is administered and allowed to remain indefinitely. For such in vitro methods, the virion can be administered to the cell by standard viral transduction methods, as known in the art and as exemplified herein. Titers of virus to administer can vary, particularly depending upon the cell type, but will be typical of that used for AAV transduction in general which is well known in the art. Additionally the titers used to transduce the particular cells in the present examples can be utilized.
[0164]The cells that can be transduced by the present recombinant AAVX virion can include any desired cell, such as the following cells and cells derived from the following tissues, human as well as other mammalian tissues, such as primate, horse, sheep, goat, pig, dog, rat, and mouse and avian species: Adipocytes, Adenocyte, Adrenal cortex, Amnion, Aorta, Ascites, Astrocyte, Bladder, Bone, Bone marrow, Brain, Breast, Bronchus, Cardiac muscle, Cecum, Cervix, Chorion, Cochlear, Colon, Conjunctiva, Connective tissue, Cornea, Dermis, Duodenum, Embryonic stem cells, Endometrium, Endothelium, Endothelial cells, Epithelial tissue, Epithelial cells, Epidermis, Esophagus, Eye, Fascia, Fibroblasts, Foreskin, Gastric, Glial cells, Glioblast, Gonad, Hepatic cells, Histocyte, Hair cells in the inner ear, auditory (organ of Corti) sensory epithelia, vestibular sensory epithelia, Ileum, Intestine, small Intestine, Jejunum, Keratinocytes, Kidney, Larynx, Leukocytes, Lipocyte, Liver, Lung, Lymph node, Lymphoblast, Lymphocytes, Macrophages, Mammary alveolar nodule, Mammary gland, Mastocyte, Maxilla, Melanocytes, Mesenchymal, Monocytes, Mouth, Myelin, Myoblasts Nervous tissue, Neuroblast, Neurons, Neuroglia, Osteoblasts, Osteogenic cells, Ovary, Palate, Pancreas, Papilloma, Peritoneum, Pituicytes, Pharynx, Placenta, Plasma cells, Pleura, Prostate, Rectum, Salivary gland, Skeletal muscle, Skin, Smooth muscle, Somatic, Spleen, Squamous, Stem cells, Stomach, Submandibular gland, Submaxillary gland, Synoviocytes, Testis, Thymus, Thyroid, Trabeculae, Trachea, Turbinate, Umbilical cord, Ureter, Uterus, and vestibular hair cells.
[0165]For example, provided herein is a method of transducing a cancer cell (e.g., lung cancer cell, non-small cell lung cancer cell), colon cell, CNS derived cell, ovarian cell, prostate cell, breast derived cell, cervical cord cell, kidney cell, salivary gland cell, or muscle cell using an AAV particle disclosed herein.
[0166]Also provided herein is a method of delivering a nucleic acid to a cancer cell (e.g., lung cancer cell, non-small cell lung cancer cell), colon cell, CNS derived cell, ovarian cell, prostate cell, breast derived cell, cervical cord cell, kidney cell, salivary gland cell, or muscle cell comprising administering to the cell an AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, or AAV-X26 particle containing a vector comprising the nucleic acid inserted between a pair of AAV inverted terminal repeats, thereby delivering the nucleic acid to the cell.
[0167]The cell of the provided methods can be an inner ear epithelial cell. Thus, the cell of the provided method can be an inner ear hair cell. The cell of the provided methods can be an inner or outer hair cell of the organ of Corti or a vestibular hair cell. The cell of the provided methods can be an inner ear supporting cell such as Hensen's, phalangal, interdental, or vestibular supporting cells.
[0168]The cell of the provided method can be an airway epithelial cell. The cell of the provided method can be a columnar, goblet or basal cell.
[0169]The cell of the provided method can be a cell of the submandibular gland. The cell of the provided method can be a ductal or acinar cell.
[0170]Provided are recombinant vectors based on AAVX. Such vectors may be useful for transducing erythroid progenitor cells or cells resistant to transduction by other serotypes of AAV. These vectors may also be useful for transducing cells with a nucleic acid of interest in order to produce cell lines that could be used to screen for agents that interact with the gene product of the nucleic acid of interest. In addition to transduction of other cell types, transduction of erythroid cells would be useful for the treatment of cancer and genetic diseases which can be corrected by bone marrow transplants using matched donors. Some examples of this type of treatment include, but are not limited to, the introduction of a therapeutic gene such as genes encoding interferons, interleukins, tumor necrosis factors, adenosine deaminase, cellular growth factors such as lymphokines, blood coagulation factors such as factor VIII and IX, cholesterol metabolism uptake and transport protein such as EpoE and LDL receptor, and antisense sequences to inhibit viral replication of, for example, hepatitis or HIV.
[0171]Provided is a vector, comprising the AAVX virus as well as AAVX viral particles. While AAVX is similar to AAV1-11, the viruses are found herein to be physically and genetically distinct. These differences endow AAVX with some unique advantages, which better suit it as a vector for gene therapy.
[0172]Furthermore, as shown herein, AAVX capsid protein is distinct from AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, or BAAV capsid proteins and exhibits different tissue tropism. AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, BAAV, and AAVX likely utilize distinct cellular receptors. AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, BAAV, and AAVX are serologically distinct and humans are not reported to have neutralizing antibodies to AAVX, thus in a gene therapy or gene transfer application, AAVX would allow for transduction of a patient who already possess neutralizing antibodies to AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, or BAAV either as a result of natural immunological defense or from prior exposure to AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, or BAAV vectors.
[0173]Each of the herein disclosed AAVX vectors, or vectors comprising AAVX capsid proteins, can have unique or distinct tissue tropism. As shown in FIG. 11, transduction profiles of AAV-X1, AAV-X5, AAV-X25 and AAV-X26 were different from that of known AAVs. AAVX1 and AAVX25 utilize α 2-3 link sialic acid as a cell attachment factor, whereas AAVX26 does not utilize sialic acid in the transduction process. Further, AAV12 (AAVX26) can be used to transduce cells lacking cell surface heparan sulfate or glycosphingolipids. Instead, mannosamine can be part of the AAV12 receptor or attachment.
[0174]As disclosed herein, cells that can be transduced by AAV12 include, but are not limited to, non-small cell lung cancer cells (e.g., NSCLC), (e.g., A549, EKVX, NC1-H226), colon cells (e.g., HCT-15), CNS derived cells (e.g., SF-268 and SF295), ovarian cells (e.g., IGROV1), prostate cells (e.g., PC-3), breast derived cells (e.g., T-47D), kidney cells (e.g., 293T), salivary glands (e.g., ductal cells), muscle cells.
[0175]Further, many of the AAV contaminated simian adenoviruses were originally isolated from pooled primary kidney cell cultures (Hull et al. 1957, 1956, 1958) originally established as part of a vaccine development program (VR-195, VR-197, VR-198, VR-200, VR-202, VR-209 and VR-353), but also from rectal swabs (VR-204), cervical cord (VR-355) and CNS cultures (VR-207) demonstrating that the tropism of the AAVs isolated from these adenovirus stocks include cells of kidney, cervical cord and CNS origin.
[0176]Thus, provided herein is a method of transducing a cancer cell (e.g., lung cancer cell, non-small cell lung cancer cell), colon cell, CNS derived cell, ovarian cell, prostate cell, breast derived cell, cervical cord cell, kidney cell, salivary gland cell, or muscle cell using an AAV particle disclosed herein.
[0177]Thus, provided herein is a method of delivering a nucleic acid to a cancer cell (e.g., lung cancer cell, non-small cell lung cancer cell) comprising administering to the cell an AAV12 particle containing a vector comprising the nucleic acid inserted between a pair of AAV inverted terminal repeats, thereby delivering the nucleic acid to the cell.
[0178]Thus, provided herein is a method of delivering a nucleic acid to a colon cell comprising administering to the cell an AAV12 particle containing a vector comprising the nucleic acid inserted between a pair of AAV inverted terminal repeats, thereby delivering the nucleic acid to the cell.
[0179]Thus, provided herein is a method of delivering a nucleic acid to a CNS derived cell comprising administering to the cell an AAV12 particle containing a vector comprising the nucleic acid inserted between a pair of AAV inverted terminal repeats, thereby delivering the nucleic acid to the cell.
[0180]Thus, provided herein is a method of delivering a nucleic acid to an ovarian cell comprising administering to the cell an AAV12 particle containing a vector comprising the nucleic acid inserted between a pair of AAV inverted terminal repeats, thereby delivering the nucleic acid to the cell.
[0181]Thus, provided herein is a method of delivering a nucleic acid to a prostate cell comprising administering to the cell an AAV12 particle containing a vector comprising the nucleic acid inserted between a pair of AAV inverted terminal repeats, thereby delivering the nucleic acid to the cell.
[0182]Thus, provided herein is a method of delivering a nucleic acid to a breast derived cell comprising administering to the cell an AAV12 particle containing a vector comprising the nucleic acid inserted between a pair of AAV inverted terminal repeats, thereby delivering the nucleic acid to the cell.
[0183]Thus, provided herein is a method of delivering a nucleic acid to a cervical cord cell comprising administering to the cell an AAV12 particle containing a vector comprising the nucleic acid inserted between a pair of AAV inverted terminal repeats, thereby delivering the nucleic acid to the cell.
[0184]Thus, provided herein is a method of delivering a nucleic acid to a kidney cell comprising administering to the cell an AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, or AAV-X26 particle containing a vector comprising the nucleic acid inserted between a pair of AAV inverted terminal repeats, thereby delivering the nucleic acid to the cell.
[0185]Thus, provided herein is a method of delivering a nucleic acid to a salivary gland cell comprising administering to the cell an AAV12 particle containing a vector comprising the nucleic acid inserted between a pair of AAV inverted terminal repeats, thereby delivering the nucleic acid to the cell.
[0186]Thus, provided herein is a method of delivering a nucleic acid to a muscle cell comprising administering to the cell an AAV12 particle containing a vector comprising the nucleic acid inserted between a pair of AAV inverted terminal repeats, thereby delivering the nucleic acid to the cell.
Vector System
[0187]Provided herein is a vector system for producing infectious virus particles having a characteristic of AAVX. As used herein, a “vector system” is a combination of one or more vectors that, when added to an appropriate cell system, can produce a recombinant AAVX virion, as provided herein. Thus, the provided vector system comprises at least one vector comprising a nucleic acid selected from the group consisting of a pair of AAV ITRs, a nucleic acid encoding an AAV capsid protein, and a nucleic acid encoding an AAV Rep protein. In addition, it is understood that an AAV vector system comprises at least one adenovirus helper plasmid.
[0188]The vector system can comprise one or more unique AAV vectors. Thus, the vector system can comprise, for example, 1, 2, 3, 4, 5, or 6 unique AAV vectors. In a three-vector system, the first AAV vector can comprise a nucleic acid encoding an AAV capsid protein, the second AAV vector can comprise a nucleic acid encoding an AAV Rep protein, and the third AAV vector can comprise a pair of AAV ITRs (Table 3). It is understood that Rep and Cap sequences can be efficiently combined in the same vector. Thus, in an example of a two-vector vector system, the first AAV vector can comprise a nucleic acid encoding an AAV capsid protein and a nucleic acid encoding an AAV Rep protein and the second AAV vector can comprise a pair of AAV ITRs (Table 3). It is understood that at least one AAV vector in the provided AAVX vector system comprises an AAVX capsid, Rep or ITR (Table 3).
[0189]Thus, provided is an AAV vector system, wherein the first vector can comprise a nucleic acid encoding an AAVX capsid protein and the second vector can comprise a pair of AAV ITRs. The AAV ITRs of the second vector can be a pair of AAV1 ITRs. The AAV inverted terminal repeats can be a pair of AAV2 ITRs. The AAV ITRs can be a pair of AAV3 ITRs. The AAV ITRs can be a pair of AAV4 ITRs. The AAV ITRs can be a pair of AAV5 ITRs. The AAV ITRs can be a pair of AAV6 ITRs. The AAV ITRs can be a pair of AAV7 ITRs. The AAV ITRs can be a pair of AAV8 ITRs. The AAV ITRs can be a pair of AAV9 ITRs. The AAV ITRs can be a pair of AAV 10 ITRs. The AAV ITRs can be a pair of AAV 11 ITRs. The AAV ITRs can be a pair of AAAV ITRs. The AAV ITRs can be a pair of BAAV ITRs. The AAV ITRs can be a pair of AAVX ITRs.
[0190]Also provided is an AAV vector system, wherein the first vector can comprise a nucleic acid encoding an AAV capsid protein and the second vector can comprise a pair of AAVX ITRs. The capsid protein can be an AAV1 capsid protein. The capsid protein can be an AAV2 capsid protein. The capsid protein can be an AAV3 capsid protein. The capsid protein can be an AAV4 capsid protein. The capsid protein can be an AAV5 capsid protein. The capsid protein can be an AAV6 capsid protein. The capsid protein can be an AAAV Rep protein. The capsid protein can be a BAAV Rep protein. The capsid protein can be an AAVX Rep protein.
[0191]In either of the above vector systems, the first vector or a third vector can further comprise a nucleic acid encoding an AAV Rep protein. The AAV Rep protein can be AAV1 Rep protein. The AAV Rep protein can be AAV2 Rep protein. The AAV Rep protein can be AAV3 Rep protein. The AAV Rep protein can be AAV4 Rep protein. The AAV Rep protein can be AAV5 Rep protein. The AAV Rep protein can be AAV6 Rep protein. The AAV Rep protein can be AAV7 Rep protein. The AAV Rep protein can be AAV8 Rep protein. The AAV Rep protein can be AAV9 Rep protein. The AAV Rep protein can be AAV10 Rep protein. The AAV Rep protein can be AAV11 Rep protein. The AAV Rep protein can be AAAV Rep protein. The AAV Rep protein can be BAAV Rep protein. The AAV Rep protein can be AAVX Rep protein.
[0192]Also provided is an AAV vector system, wherein the first vector can comprise a nucleic acid encoding an AAVX Rep protein and the second vector can comprise a pair of AAV ITRs. The AAV ITRs of the second vector can be a pair of AAV1 ITRs. The AAV inverted terminal repeats can be a pair of AAV2 ITRs. The AAV ITRs can be a pair of AAV3 ITRs. The AAV ITRs can be a pair of AAV4 ITRs. The AAV ITRs can be a pair of AAV5 ITRs. The AAV ITRs can be a pair of AAV6 ITRs. The AAV ITRs can be a pair of AAV7 ITRs. The AAV ITRs can be a pair of AAV8 ITRs. The AAV ITRs can be a pair of AAV9 ITRs. The AAV ITRs can be a pair of AAV10 ITRs. The AAV ITRs can be a pair of AAV 11 ITRs. The AAV ITRs can be a pair of AAAV ITRs. The AAV ITRs can be a pair of BAAV ITRs. The AAV ITRs can be a pair of AAVX ITRs.
[0193]The first vector or a third vector can further comprise a nucleic acid encoding an AAV Capsid protein. The capsid protein can be an AAV1 capsid protein. The capsid protein can be an AAV2 capsid protein. The capsid protein can be an AAV3 capsid protein. The capsid protein can be an AAV4 capsid protein. The capsid protein can be an AAV5 capsid protein. The capsid protein can be an AAV6 capsid protein. The capsid protein can be an AAAV Rep protein. The capsid protein can be a BAAV Rep protein. The capsid protein can be an AAVX Rep protein.
[0000] |
| AAVX Vector Systems |
| First Vector | Second Vector | Third Vector |
| |
| AAVX capsid + AAV Rep | AAV ITR | — |
| AAV capsid + AAV Rep | AAVX ITR | — |
| AAV capsid + AAVX Rep | AAV ITR |
| AAVX capsid | AAV ITR | AAV Rep |
| AAV capsid | AAVX ITR | AAV Rep |
| AAV capsid | AAV ITR | AAVX Rep |
| |
| “AAV” includes: AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAAV, BAAV, AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, or AAV-X26. |
| “AAVX” includes: AAV-X1, AAV-X1b, AAV-X5, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, or AAV-X26. |
[0194]In either of the above vector systems, the second vector comprising a pair of AAV ITRs can further comprise a promoter between the ITRs. The promoter can be AAV2 p5 promoter. The promoter can be AAV5 p5 promoter. The promoter can be AAVX p5 promoter. Furthermore, smaller fragments of p5 promoter that retain promoter activity can readily be determined by standard procedures including, for example, constructing a series of deletions in the p5 promoter, linking the deletion to a reporter gene, and determining whether the reporter gene is expressed, i.e., transcribed and/or translated. The promoter can be the AAVX p19 promoter. The promoter can be the AAVX p40 promoter. The promoter can be a promoter of any of the AAV serotypes. The promoter can be a constitutive promoter. Thus, the promoter can be CMV. The promoter can be RSV. The promoter can be LTR. The promoter can be eFl. The promoter can be beta actin promoter. The promoter can be a tissue specific promoter. The promoter can be an inducible promoter. The promoter can further be functionally linked to an exogenous nucleic acid.
[0195]Further provided is any of the disclosed vectors of the vector systems encapsidated into an AAV particle. The AAV particle can be an AAV1 virus particle comprising at least one AAV1 capsid protein. The AAV particle can be an AAV2 virus particle comprising at least one AAV2 capsid protein. The AAV particle can be an AAV3 virus particle comprising at least one AAV3 capsid protein. The AAV particle can be an AAV4 virus particle comprising at least one AAV4 capsid protein. The AAV particle can be an AAV5 virus particle comprising at least one AAV5 capsid protein. The AAV particle can be an AAV6 virus particle comprising at least one AAV6 capsid protein. The AAV particle can be an AAV7 virus particle comprising at least one AAV7 capsid protein. The AAV particle can be an AAV8 virus particle comprising at least one AAV8 capsid protein. The AAV particle can be an AAV9 virus particle comprising at least one AAV9 capsid protein. The AAV particle can be an AAV10 virus particle comprising at least one AAV10 capsid protein. The AAV particle can be an AAV 11 virus particle comprising at least one AAV 11 capsid protein. The AAV particle can be an AAAV virus particle comprising at least one AAAV capsid protein. The AAV particle can be a BAAV virus particle comprising at least one BAAV capsid protein. The AAV particle can be an AAVX virus particle comprising at least one AAVX capsid protein. The AAV particle can be a chimeric capsid virus particle (described above) comprising a capsid protein from more than one serotype of AAV.
EXAMPLES
[0196]It is understood that the disclosed method and compositions are not limited to the particular methodology, protocols, and reagents described as these may vary. It is also to be understood that the terminology used herein is for the purpose of describing particular embodiments only, and is not intended to limit the scope of the present invention which will be limited only by the appended claims.
Example 1
Identification and Characterization of Novel AAV Isolates in ATCC Virus Stocks
[0197]Materials and Methods
[0198]Cell culture and virus propagation: 293T and COS cells were maintained in DMEM, supplemented with 10% FBS, 2 mM L-glutamine, 100 U/ml penicillin, and 0.1 mg/ml streptomycin or RPMI with 5% FBS. SF295, MCF7, EKVX, Igrov1, CAKI, Ovcar 5 cells were cultured in RPMI supplemented with 5% FBS, 2 mM L-glutamine, 100 U/ml penicillin, and 0.1 mg/ml streptomycin. Cells were maintained at 37° C. in a 5% CO2humidified atmosphere.
[0199]Screening for novel AAVs: 109 virus samples, obtained from the ATCC (American Type Culture Collection) as lysate of infected cells, were analyzed for the presence of AAV DNA by PCR as described earlier (Katano, H., et al. 2004. Biotechniques 36:676-80). Briefly, low molecular weight DNA was purified with High Pure Viral Nucleic Acid Kit (Roche). These DNA samples were assayed for AAV contamination by PCR using the GC Rich PCR Kit (Roche). This method detects the presence of AAV DNA by PCR using degenerative PCR primers, which were shown to amplify a fragment containing sequences of the rep and vp ORF of all known AAV serotypes. PCR using DNA isolated from ATCC VR-195, VR-197, VR-198, VR-200, VR-202, VR-204, VR-207, VR-209, VR-353, VR-355, VR-942, VR-943 as template resulted in the generation of a 1.4 kb amplification product, which was subsequently cloned using the TOPO TA Cloning KIT (Invitrogen) and sequenced with an ABI Prism 3100 Genetic Analyzer (ABI) and FS dye-terminator chemistry (ABI). The obtained sequences showed homology to AAV6 rep ORF and cap ORF but were not identical to any known AAV.
[0200]Viral DNA isolation, cloning and sequencing: The rep and cap ORF of the new AAVs was PCR amplified and subcloned. Viral DNA that was isolated from VR-195, VR-197, VR-198, VR-200, VR-202, VR-204, VR-207, VR-209, VR-353, VR-355, VR-942, VR-943 was PCR amplified with primers:
[0000] | AAV1-4 225(+): | |
| GCGACAKTTTGCGACACCAYGTGG | (SEQ ID NO: 31) |
| and |
| |
| UNI-NC: |
| CCANNNGGAATCGCAATGCCAAT, | (SEQ ID NO: 32) |
| or |
| |
| UNIC: |
| ATGNTNATNTGGTGGGAGGAGGG | (SEQ ID NO: 33) |
| and |
| |
| AAV1-4 polyA4400(−): |
| CGAATNAAMCGGTTTATTGATTAAC, | (SEQ ID NO: 34) |
| or |
| |
| AAV 4500 (+): |
| CAATAAACCGkkTnATTCGTkTCAGT | (SEQ ID NO: 38) |
| and |
| |
| AAV 450 (−): |
| ACANNWGAGTCAGAAATKCCNGGCAG | (SEQ ID NO: 39) |
(N can be A, C, G, or T; K can be G or T; Y can be C or T; M can be A or C)
[0201]to amplify the Rep ORF, capsid ORF, and ITR(3′-AAVX terminus, ITR and 3′-terminus of circular or concatermerized AAVs), respectively. The PCR products were subcloned using the TOPO TA Cloning KIT (Invitrogen) and at least three clones of each isolate were sequenced with an ABI Prism 3100 Genetic Analyzer (ABI) and FS dye-terminator chemistry (ABI). The obtained sequences showed homology to AAV1 and AAV6 but were not identical to any known AAV. As shown in Table 5, there are three naming conventions used herein. For example, the newly identified AAVs were named according to the ATCC adenovirus strains from which they were discovered (e.g., AAV(VR-943)). Also, each unique AAV isolate was assigned a temporary identifier (e.g., AAV-X26). Finally, . . . (e.g., AAV-12).
[0202]Identical AAV sequences were detected in VR-195, VR-197, VR-198 and VR-202 and named AAV-X1. VR-195 also contained a homolog of AAV-X1 termed AAV-X1b.
[0203]AAV detected in VR-200 was named AAV-X19. AAV detected in VR-204 was named AAV-X21. AAV detected in VR-207 was named AAV-X22. AAV detected in VR-209 was named AAV-X23. AAV detected in VR-353 was named AAV-X24. AAV detected in VR-355 was named AAV-X25. AAV detected in VR-942 was named AAV-X5. AAV detected in VR-943 was named AAV-X26. (Table 5).
[0204]Sequence analysis: DNA and protein sequence alignments were performed using the Clustal W multiple sequence alignment tool of the Biology Workbench web based software (SDSC), MacVector 7 (Oxford Molecular). The sequences amplified from ATCC virus stocks were compared to sequences in GenBank using BLAST (Basic Local Alignment Search Tool at http://www.ncbi.nlm.nih.gov/BLAST/). DNA alignments were performed using the ClustalW multiple sequence alignment tool of the Biology Workbench web based software at http://seqtool.sdsc.edu (SDSC) and MacVector 7 (Accelrys, Burlington, Mass.). Divergent amino acids in VP1 were mapped on the capsid by superimposing the VP1 sequence onto a pseudoatomic structure for AAV-6. The AAV6 crystal structure was predicted by remodeling the AAV2 capsid structure, obtained from the Virus Particle Explorer, at the Swiss-Model server with the AAV6 capsid sequence.
[0205]AAV-X17 partial ITR sequence was 98% identical to corresponding AAV2 ITR sequence. AAV-X22 partial ITR sequence was 99% identical to corresponding AAV2 ITR sequence. AAV-X25 partial ITR sequence was 96% identical to corresponding AAV1 ITR sequence. AAV-X26 was only about 80% identical to corresponding AAV2 ITR sequence. However, the TRS signal (e.g., aa 176-181 of SEQ ID NO:40) and Rep Binding site (e.g., aa 195-210 of SEQ ID NO:40) were conserved in AAV-X26 ITR as compared to the AAV2 ITR.
[0206]Generation of recombinant virus: Recombinant AAV-X1, AAV-X1b, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, AAV-X5, and AAV-X26 were generated by transfecting 293 T cells with AAV2 vector plasmid (AAV2-NLS-GFP) consisting of an GFP expression cassette flanked by AAV-2 ITRs and AAV-X1, AAV-X1b, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, AAV-X5, or AAV-X26 packaging (subcloned UNI-C/AAV1-4 polyA4400 (−) PCR fragment), an AAV-2 like Rep expression plasmid, AAV-X26_Rep, (subcloned AAV-X26 Rep ORF, AAV1-4 225 (+) and UNI-NC PCR fragment), and Ad helper plasmids, 449B, which provided the essential adenovirus functions that are required for AAV replication (Smith, R. H., et al. (2002). Biotechniques 33(1): 204-6, 208, 210-1). Two confluent T175 flasks of 293T cells were harvested, resuspended in 100 ml DMEM 10% FCS, seeded in five 150 mm plates and incubated at 37° C., 5% CO2until cells are 80% confluent (typically 48 h). Cells were transfected with 15 μg pAAV2-NLS-GFP, 10 μg AAV-X1, AAV-X1b, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, AAV-X5, or AAV-X26 packaging plasmid, 5 μg AAV-X26 Rep and 30 μg p449B. 48 h after transfection, cells were harvested, washed with PBS and resuspended in 11 ml TD buffer (0.14 M NaCl, 5.0 mM KCl, 0.7 mM K2HPO4, 25.0 mM Tris, pH7.4. Cells were lyzed by 3 freeze thaw cycles and incubated for 30 minutes at 37° C. after adding benzonase to a final concentration of 20 U/ml sodium deoxycholate (final concentration of 0.5%). After adding 0.55 g/ml CsCl, the lysate was fractionated using density gradient centrifugation in a SW41 rotor for 48 h at 38,000 rpm. The gradients were harvested in 0.5 ml aliquots. Aliquots were assayed for infectivity and DNase-resistant genome copy titers of the vector preparations were determined by quantitative real-time PCR using the TAQMAN system (Applied Biosystems, Inc. (ABI), Foster City, Calif.) with probes specific to the CMV promoter.
[0207]Recombinant virus was successfully generated using each of the AAVX packaging plasmids with AAV2 ITR vector plasmids and AAV-X26 rep plasmids. Recombinant virus was also successfully generated using Rep plasmids derived from each of the AAVX viruses in combination with AAV2 ITR vector plasmids. There is therefore sufficient homology between AAVX and AAV2 Rep and ITR sequences that they can be used interchangeably in the herein provided vectors and vector systems.
[0208]Neutralization assay: Exponentially growing COS cells were plated at a density of 5×103/well in a flat-bottom 96-well plate. Twenty-four hours after seeding, cells were incubated for 60 min with 2×106 rAAV particles that were pre-incubated with serial dilutions of pooled human IgGs (Immune Globuline Intravenous, 10%, Gamunex, BAYER, Leverkusen, Germany). Twenty-four hours after infection, cells were analyzed for GFP expression with the Guava PCA-96 (Guava Technologies, Hayward, Calif.) fluorescent cell counter. GFP expression was used as a surrogate marker for transduction efficiency.
[0209]Determination of tissue tropism: Transduction efficiency of recombinant AAV-X1, AAV-X1b, AAV-X19, AAV-X21, AAV-X22, AAV-X23, AAV-X24, AAV-X25, AAV-X5, or AAV-X26 vector containing an expression cassette for GFP was analyzed in cancer cell lines. Cells were infected with dilutions ranging from 106to 109particles/well. 24-48 h after infection, cells were analyzed for GFP expression by flow cytometry.
[0210]Neuraminidase treatment: To analyze if sialic acid is required for transduction of AAV-X1 and AAV-X25, neuraminidases were used to digest cell surface sialic acid before infection. Cos cells were seeded at 5,000 cells/well in a 96 well plate. 24 h after seeding, cells were incubated for 60 min with a broad spectrum neuraminidase from Arthrobacter ureafaciens (0.05-1 U/ml) or a 2-3 linkage specific neuraminidase from S. pneumoniae (0.04-0.2 U/ml) (Calbiochem, La Jolla, Calif.). Cells were washed and transduced with a multiplicity of infection (MOI) of 500 with recombinant AAV6, AAV-X1 or AAV-X25 particles expressing GFP. Cells were analyzed for GFP expression by flow cytometry with the Guava PCA-96 (Guava Technologies) 24 h after transduction.
[0211]Binding assay: COS cells were seeded at 5000 cells/well in a 96-well plate. Forty-eight hours after seeding, cells were incubated for 60 min at 37° C. with neuraminidases from A. ureafaciens (0.1 U/ml), S. pneumoniae (0.08 U/ml) (Calbiochem) or mock. Cells were then chilled for 30 min at 4° C. and then incubated for 60 min at 4° C. with 5×107recombinant AAV6, AAV(VR-195), or AAV(VR-355) particles expressing GFP. Cells were then washed twice with cold medium and phosphate buffered saline (PBS) and iysed in 50 μl PCRnGo buffer (Pierce). Copy numbers of cell-associated vector genomes in the cell lysates were determined by quantitative real-time PCR using the TAQMAN system (Applied Biosystems) with probes specific to the CMV promoter.
[0212]Lectin competition: COS cells were seeded at 5,000-10,000 cells/well in a 96 well plate. 16-24 h after seeding, cells were precubated for 30 min at 4° C. with 100 μg/ml of either ConA, MalII, LCA, ECL, WGA, UEA I, DBA, PNA, SBA, GSL I, PSA, LCA, PHA-E, PHA-L, SJA, succinylated WGA. Subsequently, cells were washed and transduced for 60 min at 4° C. with 3000 transducing units or 108recombinant AAV-2, AAV-4, AAV-5, AAV-6, BAAV, AAV-X1, AAV-X5, AAV-X25 or AAV-X26 particles expressing GFP in 50 μl medium supplemented with lectins (100 μg/ml). 24 h after transduction, cells were analyzed for GFP expression by flow cytometry.
[0213]Sugar competition: COS cells were seeded at 5,000 cells/well in a 96 well plate. 24 h after seeding, cells were precubated for 30 min at 4° C. before being transduced for 60 min at 4° C. with 108recombinant AAV-2, AAV-4, AAV-5, AAV-6, BAAV, AAV-X1, AAV-X5, AAV-X25 or AAV-X26 particles expressing GFP in 50 μl medium supplemented with various sugars (see FIG. 9). 24 h after transduction, cells were analyzed for GFP expression by flow cytometry.
[0214]Heparin competition assay: COS cells were plated at a density of 5×103/well in a flat-bottom 96-well plate. After 24 h, 107particles rAAV6-NLS-GFP, rAAV(VR-195)-NLS-GFP and rAAV(VR-355)-NLSGFP were preincubated for 1 h at room temperature in medium containing 0 to 2000 μg/ml heparin (Sigma, St. Louis, Mo.). Cells were then transduced for 1 h at 37° C. with this preincubation mixture, washed with medium and incubated for 24 h. GFP expression was detected with a fluorescent cell counter (Guava Technologies).
[0215]NaCl competition assay: COS cells were seeded at 5,000 cells/well in a 96 well plate. 24 h after seeding, were transduced for 60 min at 37° C. with MOI of 500 or 108recombinant AAV-2, AAV-4, AAV-5, AAV-6, BAAV, AAV-X1, AAV-X5, AAV-X25 or AAV-X26 particles expressing GFP in 50 μl medium supplemented with NaCl to 150 mM, 300 mM, and 450 mM final NaCl concentration. 24 h after transduction, cells were analyzed for GFP expression by flow cytometry/fluorescent cell counter (Guava Technologies).
[0216]Viral cross-competition: COS cells were plated at a density of 5×103/well in a flat-bottom 96-well plate. After 24 h, cells were preincubated for 1 h at 37° C. with increasing titers of rAAV6-lacZ, an AAV6 derived vector expressing a nuclear localized β-galactosidase, ranging from 0 to 2.6×109particles/well. The cells were then washed with medium and transduced for 45 min with 2×106particles of rAAV2-NLS-GFP, rAAV6-NLS-GFP, rAAV(VR-195)-NLS-GFP, or rAAV(VR-355)-NLS-GFP. GFP expression was analyzed 48 h after transduction by flow cytometry.
[0218]Identification of AAV contaminations in ATCC virus isolates: Viral stocks, supplied by the ATCC were analyzed for the presence of AAV DNA by a PCR based assay as described earlier (Katano, H., et al. 2004). PCR primers in this study were designed to bind to highly conserved regions in the rep and cap ORFs resulting in amplification of a 1.5 kb fragment spanning from nucleotide 1437 to 2904 relative to the AAV-2 genome. This method is highly sensitive and can detect as few as 10 copies of viral DNA/μL of sample (Katano, H., et al. 2004). AAV DNA was detected in 13/137 samples analyzed (Table 4). AAV contaminations were detected in 26% of adenovirus isolates. Interestingly, no AAV was detected in herpesvirus, retrovirus, coronavirus, orthomyxovirus, poxvirus, or reovirus stocks. Many of the AAV contaminated simian adenoviruses were originally isolated from pooled primary kidney cell cultures (Hull, R. N., et al. 1957; Hull, R. N., et al. 1958; Hull, R. N., et al. 1956) originally established as part of a vaccine development program (VR-195, VR-197, VR-198, VR-200, VR-202, VR-209, and VR-353), but also from rectal swabs (VR-204), cervical cord (VR-355), and CNS cultures (VR-207). AAV was also detected in stocks of human adenovirus type 9 (VR-10) as well as in bovine adenovirus type 1 (VR-313) and type 2 (VR-314). Ten of theses isolates have high similarity to AAV1 and AAV6 (>98%), while AAV-X5 isolated from VR-942 and AAV-X26 isolated from VR-943 showed highest homology in the capsid protein VP-1 to AAV-3B (93%) and AAV-11 (83%) respectively.
[0219]Construction of packaging plasmids and generation of recombinant virus: The entire coding region for Rep and Cap of the AAV contaminations detected in VR-195 and VR-355 termed AAV(VR-195) and AAV(VR-355), respectively, as well as the capsid ORF under control of the viral P40 promoter from the adenovirus isolates VR-10, VR-195, VR-197, VR-198, VR-200, VR-202, VR-209, VR-353, VR-204, VR-355, and VR-207 were PCR amplified and subcloned to generate packaging plasmids.
[0000] |
| Screening of viral samples for the presence of AAV |
| Family | # tested | # AAV positive | frequency (%) |
|
| Adenovirida | 53 | 13 | 26% |
| primate Adenovirida | 16 | 13 | 81% |
| Herpesviridae | 43 | 0 | 0 |
| Coronaviridae | 9 | 0 | 0 |
| Retrovirida | 15 | 0 | 0 |
| Reoviridae | 4 | 0 | 0 |
| Orthomyxovirida | 4 | 0 | 0 |
|
[0220]Recombinant AAV was produced by cotransfecting plasmids encoding the capsid of the novel AAVs, an AAV type 2 Rep expression plasmid, an AAV-2 vector plasmid, encoding a nuclear localized GFP together with an adenovirus helper plasmid. Recombinant viruses were then assayed for transduction activity. Capsid plasmids encoding functional VP proteins were sequenced on both strands. Several clones of each isolate were analyzed. GenBank accession numbers are as follows: AAV(VR-195): DQ180604 (SEQ ID NO:1) and AAV(VR-355): DQ180605 (SEQ ID NO:9).
[0221]The evolutionary relationship among mammalian AAVs and the AAV contaminants detected in human and non-human primate adenovirus stocks was analyzed by ClustalW alignments of VP1 amino acid sequences and plotted as a rooted phylogenetic tree (FIG. 1). All AAVs detected in simian adenovirus stocks displayed at least 96% homology on the DNA level and 98% identity in the capsid amino acid sequence to either AAV1, AAV6, or to each other. The amino acid sequence of VP1 encoded by AAV contaminants in VR-197, VR-198, and VR-202 were identical to each other but distinct from AAV1 and AAV6. The VP1 sequence of the only positive human adenovirus stock, VR-10 (adenovirus type 9), was greater than 99.9% identical to AAV2.
[0222]AAV-X1(97%, 99%), AAV-X1B (96%, 98%), AAV-X19 (97%, 99%), AAV-X21(97%, 99%), AAV-X22 (97%, 99%), AAV-X23(97%, 98%), AAV-X24(97%, 97%), and AAV-X25 (97%, 98%) share 96-99% sequence identity to AAV6 or AAV1 at the DNA level (based on a 3 kb partial genome fragment) (see FIG. 1). Numbers in brackets indicate identities to AAV6 and AAV1. The amino acid sequence of AAV-X5 capsid protein shows highest homology to AAV-3B (93%), and AAV-X26 capsid protein shows highest homology to AAV-11 (83%). AAV-X5 and AAV-X26 DNA sequences show highest homology to AAV-3 (88%) and AAV-11 (79%) respectively (based on a 3 kb partial genome fragment) (see FIG. 1).
[0223]Mapping of sequence variation: AAV contaminations, which were greater than 98% identical to AAV6 in the amino acid sequence of the capsid protein VP1, were detected in 10 adenovirus isolates. AAV-X5 isolated from VR-942 and AAV-X26 isolated from VR-943 showed homology (sequence similarity) in the capsid protein VP1 to AAV-3B (93%) and AAV-11 (83%), respectively, and only low homology to AAV6 (87% and 61%, respectively). AAV-X5 and AAV-X26 are therefore very different from the other new AAVs disclosed herein. The capsid protein VP1 of AAV(VR-195) and AAV(VR-355) differs in 7 or 6 amino acids, respectively, from that of AAV6 (Table 5). To identify the location of the amino acids that are unique for AAV(VR-195) and AAV(VR-355) within the capsid, we superimposed the AAV(VR-195) and AAV(VR-355) VP1 sequence onto a pseudoatomic structure for AAV-6. Divergent regions in the capsid proteins are located on exposed surface loops at the threefold axis of symmetry, an area of the capsid that has been associated with receptor binding (Kern, A., et al. 2003; Opie, S. R., et al. 2003). AAV(VR-195) specific amino acids of three different VP3 subunits were clustered in close proximity. AAV(VR-355) specific changes relative to AAV6 were similarly organized. Since the amino acid changes among AAV(VR-195), AAV(VR-355), and AAV6 are surface exposed, we hypothesized they may effect antigenicity or cell tropism.
[0000] |
| Novel AAVs were isolated from ATCC adenovirus samples |
| | Isolated | | |
| Name | A.K.A. | from | Classification | Family |
|
| AAV-X1, | AAV(VR-195) | VR-195 | Simian virus 1 | Adenovirida |
| AAV-X1b |
| AAV-X1 | AAV(VR-197) | VR-197 | Simian virus 15 | Adenovirida |
| AAV-X1 | AAV(VR-198) | VR-198 | Simian virus 17 | Adenovirida |
| AAV-X19 | AAV(VR-200) | VR-200 | Simian virus 23 | Adenovirida |
| AAV-X1 | AAV(VR-202) | VR-202 | Simian virus 27 | Adenovirida |
| AAV-X21 | AAV(VR-204) | VR-204 | Simian virus 31 | Adenovirida |
| AAV-X22 | AAV(VR-207) | VR-207 | Simian virus 34 | Adenovirida |
| AAV-X23 | AAV(VR-209) | VR-209 | Simian virus 37 | Adenovirida |
| AAV-X24 | AAV(VR-353) | VR-353 | Simian virus 39 | Adenovirida |
| AAV-X25 | AAV(VR-355) | VR-355 | Simian virus 38 | Adenovirida |
| AAV-X5 | AAV(VR-942) | VR-942 | Simian | Adenovirida |
| | | adenovirus 17 |
| AAV-X26 | AAV(VR-943) | VR-943 | Simian | Adenovirida |
| AAV-12 | | adenovirus 18 |
|
[0224]Immunological characterization: To test for a difference in the immunological response to the isolates, IgGs purified from pooled human serum were assayed to determine if they contained neutralizing antibodies against the recombinant AAVs and whether a difference in the neutralization activity against either AAV6, AAV(VR-195) or AAV(VR-355) existed (FIG. 2). In this assay, all three viruses displayed similar sensitivity to neutralization with the purified pooled IgGs. Thus, the changes in the capsid of AAV(VR-195) or AAV(VR-355) compared to AAV6 and do not appear to alter their sensitivity to neutralization by human serum.
[0225]Heparin competition: Heparan sulfate, a ubiquitous cell surface glycosaminoglycan, is an attachment receptor for AAV2 (Summerford, C., et al. 1998). AAV2 transduction can be inhibited by heparin, a heparan sulfate analog. Like AAV2, AAV6 can be purified using a heparin affinity column; however, transduction is not inhibited by a low heparin concentration, indicating that heparan sulfate does not act as an AAV6 receptor (Halbert, C. L., et al. 2001). To investigate the role of heparan sulfate in AAV(VR-195) or AAV(VR-355) transduction, competition transduction experiments were performed with increasing amounts of heparin. At low concentrations of heparin (25 μg/ml), AAV2 was the only rAAV that was inhibited (FIG. 3). However, at 2000 μg/ml, approximately 50% inhibition of AAV6 and AAV(VR-195) was observed. No inhibition was observed with either AAV(VR-355) or AAV5. Therefore, heparan sulfate does not appear to be a co-receptor for either AAV(VR-195) or AAV(VR-355).
[0226]AAV6 and AAV(VR-195) transduction is charge sensitive: Heparin is a highly charged molecule. The observed inhibition of AAV6 and AAV(VR-195) at high concentrations of heparin could be caused by a charge driven interaction between the viruses and heparin rather than a specific interaction. Increasing the ionic strength in the medium during transduction can minimize charge dependent interactions (Amberg, N., et al. 2002). Salt ions can bind to charged groups on the cell surface and virus capsid, thereby reducing these electrostatic interactions. To analyze if AAV6, AAV(VR-195), and AAV(VR-355) transduction is charge dependent, the ionic strength was increased during transduction by adjusting the NaCl concentration in the tissue culture medium from 150 mM up to 450 mM. Transduction of rAAV-6, rAAV(VR-195), and rAAV(VR-355) was inhibited to different amounts by increasing concentrations of NaCl (FIG. 4). While 250 mM NaCl was sufficient to inhibit rAAV6 by 50%, higher NaCl concentrations were required to inhibit AAV(VR-195) and AAV(VR-355) with IC50of 350 mM and 450 mM respectively. The relative dependence on charge follows the inhibition of these viruses by heparin and indicates that the inhibition of AAV6 and AAV(VR-195) by heparin is charge dependent.
[0227]Effect of neuraminidase treatment on rAAV transduction and virus binding: Dependent parvoviruses are also reported to use sialic acid as a co-receptor for binding and transduction (Kaludov, N., et al. 2001; Seiler, M. P., et al. 2002; Walters, R. W., et al. 2001). To analyze if sialic acid is required for transduction with AAV(VR-195) and AAV(VR-355) vectors, the effect of the removal of cell surface sialic acids was studied by neuraminidase treatment on transduction (FIG. 5A) and virus binding (FIG. 5B). Treating COS cells with a broad-spectrum neuraminidase from A. ureafaciens inhibited rAAV6, rAAV(VR-195), and rAAV(VR-355) transduction 3-5 fold. Neuraminidase isolated from S. pneumoniae will only remove ce 2-3 linked sialic acid. Treatment of the cells with this enzyme again inhibited transduction with all three rAAVs. Removal of cell surface sialic acids with neuraminidases from A. ureafaciens or S. pneumoniae also resulted in a greater than 90% reduction in cell binding of rAAV6, rAAV(VR-195) and rAAV(VR-355), indicating that these viruses utilize α 2-3 link sialic acid as an cell attachment factor.
[0228]AAV6, AAV(VR-195), and AAV(VR-355) can be distinguished based upon lectin competition: Lectins are proteins that recognize and bind oligosaccharides conjugated to proteins and lipids and can be used to block virus binding (Summerford, C., et al. 1998). Ma111, a lectin that recognizes α 2-3 linked sialic acid, inhibited transduction of rAAV-6, rAAV(VR-195), and rAAV(VR-355) (FIG. 6).
[0229]Many glycoproteins contain a core oligosaccharide structure which includes α-linked mannose. To study, if these “high-mannose” proteins play a role in AAV-X1 or AAV-X25 transduction, these proteins were blocked with ConA, a lectin that recognizes α-linked mannose. AAV-6, AAV-X1 or AAV-X25 were blocked equally by ConA, indicating that proteins with a mannose core are involved in binding and/or uptake. Lens Culinaris Agglutinin (LCA) recognizes α-linked mannose together with neighboring oligosachharides. It is less broad specific than ConA. LCA inhibited all recombinant viruses tested and had an approximately twofold higher inhibitory potential on rAAV6 than on rAAV(VR-195) or rAAV(VR355). ECL is a lectin specific toward galactose residues and had the highest binding activity toward galactosyl (β-1,4) N-acetylglucosamine. ECL blocked AAV-X1 mediated transduction less specific than AAV-6 or AAV-X25. While ECL, which recognizes α-mannose in conjugation with galactosyl (β-1,4) N-acetylglucosamine, did not inhibit rAAV(VR-195), it reduced rAAV6 and rAAV(VR-355) transduction 7 and 5-fold, respectively. STL, which recognizes Nacetylglucosamine, did not inhibit rAAV(VR-195), but reduced rAAV6 and rAAV(VR-355) transduction 30 and 12-fold, respectively. These results indicate that while all three viruses bind terminal α 2-3 linked sialic acid, the amino acid changes on the capsid surface affected the cell binding activity of each isolate.
[0230]These differences observed between AAV-6, AAV-X1 and AAV-X25 in lectin competition experiments and NaCl competition indicates utilization of different receptors or differences in receptor interaction that can result in a different cell tropism of vectors based on these isolates. The transduction efficiency of AAV-6, AAV-X1 and AAV-X25 based vectors were therefore analyzed in human cancer cell lines (FIG. 9). Each recombinant virus has a unique transduction profile indicating that they bind to different receptors or interact differently with a common receptor.
[0231]AAV-X5 isolated from VR-942 and AAV-X26 isolated from VR-943 showed homology (sequence similarity) in the capsid protein VP1 to AAV-3B (93%) and AAV-11 (83%), respectively, and only low homology to AAV6 (87% and 61%, respectively). AAV-X5 and AAV-X26 are therefore very different from the other new AAVs disclosed herein. To study the attachment factors that are involved in AAV-X5 and AAV-X26 mediated transduction, competition with various carbohydrates was analyzed (FIG. 10). Heparin is a known attachment factor/receptor for AAV2 and addition of Heparin and homolog sugars to the medium during transduction resulted in an inhibition of AAV-2 mediated gene transfer. Heparin had no effect on AAV-X5 and AAV-X26 mediated gene transfer demonstrating that extracellular heparin can not block AAV-X5 and AAV-X26 mediated gene transfer. Heparin and related carbohydrates do not appear to have receptor function for these viruses.
[0232]AAV-X5 and AAV-X26 therefore utilize unique uptake pathways/receptors that are different from all other AAVs analyzed thus far. Transduction efficiencies in four cancer cell lines were therefore examined (FIG. 11). AAV-X5 and AAV-X26 have unique transduction properties and tropism. Thus, the application range and potential use of recombinant vectors based on AAV-X5 and AAV-X26 are unique and different from other AAVs described thus far.
[0233]rAAV6, rAAV(VR-195), and MAV(VR-355) differ in their cell tropism: As a result of the few sequence changes on the surface of the rAAV(VR-195) and rAAV(VR-355) capsid, each virus appears to exhibit different biological characteristics from each other and from rAAV6. To analyze if this difference also results in a change in tropism or transduction activity, six human cancer cell lines and African green monkey kidney cells, COS, were transduced with recombinant vectors based on rAAV6, rAAV(VR-195), and rAAV(VR-355) (FIG. 7). AAV6 and AAV(VR-195) transduced COS cells, the non-small cell lung cancer cell line EKVX, ovarian IGROV1, and renal CAKI cells with similar efficiency. However, the transduction rates of AAV(VR-195) on ovarian Ovcar5 cells and the CNS derived SF295 cell line were approximately 4 times lower compared to AAV6. AAV(VR-355) demonstrated efficient gene transfer in COS and EKVX cells, but transduction of Igrov1, CAKI, Ovcar5 and SF295 was 10 to 17-fold lower than for AAV6. The different transduction efficiencies of rAAV6, rAAV(VR-195), and rAAV(VR-355) indicate that each isolate may have a distinct cell tropism which could be the result of utilization distinct receptors for attachment or internalization or have different affinities for a common receptor.
[0234]rAAV6 competition: rAAV6, rAAV(VR-195), and rAAV(VR-355) require 2-3 linked sialic acid for cell attachment and transduction but differ in their charge dependency, sensitivity to lectin competition, and transduction activity on a panel of cells. To investigate if these viruses use distinct receptors, competition experiments were used to assay for change in transduction between AAV6 and the other isolates. COS cells were pre-incubated with increasing doses of rAAV6-lacZ followed by transduction with identical particle titers of either rAAV2, rAAV6, rAAV(VR-195), or rAAV(VR-355) expressing GFP. Changes in GFP expression were detected by flow cytometry (FIG. 8). rAAV6-LacZ competition had the greatest effect on rAAV6-NLS-GFP transduction. Fifty percent inhibition of rAAV6-NLS-GFP transduction was observed at approximately 60-fold excess of the competitor, whereas a 220-fold excess was required for the same level of inhibition of rAAV(VR-195) or rAAV(VR-355). Fifty percent inhibition of rAAV2-NLS-GFP transduction required greater than 250-fold particle excess. The stronger inhibition of rAAV6-NLS-GFP by rAAV6-lacZ compared with rAAV(VR-195) or rAAV(VR-355) suggests, that while rAAV6, rAAV(VR-195) and rAAV(VR-355) share a common attachment factor and potentially a common receptors, differences in the attachment factor and receptor interaction exist.
Example 2
Identification and Characterization of AAV12
[0235]Materials And Methods
[0236]Cells culture and virus: African green monkey kidney COS cells, obtained from the American Type Culture Collection (ATCC) (Manassas, Va.), were cultured in RPMI-1640 medium (Biosource, Camarillo, Calif.), supplemented with 5% fetal bovine serum (Hyclone, Logan, Utah), 2 mM L-glutamine, 100 U of penicillin/ml, and 0.1 mg of streptomycin/ml (Invitrogen, Carlsbad, Calif.). Cells were maintained at 37° C. under a 5% CO2humidified atmosphere. Simian Virus 18 isolate, VR-943, was obtained from the ATCC as crude lysate of virus-infected cells.
[0237]Subcloning of the AAV12 rep and cap gene: The complete coding region of AAV12 rep and cap were PCR amplified and subcloned. DNA was isolated from lysate of Simian Virus 18 infected cells with the QIAprep Spin Miniprep kit (Qiagen, Valencia, Calif.).The rep open reading frame (ORF) was PCR amplified from this DNA with the primers AAV225(+):GCGACAKTTTGCGACACCAYGTGG (SEQ ID NO:44) and UNI-NC: CCANNNGGAATCGCAATGCCAAT (SEQ ID NO:45). AAV12 cap was amplified with the primers UNI-C: 5′-ATGNTNATNTGGTGGGAGGAGGG-3′ (SEQ ID NO:46) and AAV1-4 polyA4400(−):5′-CGAATNAAMCGGTTTATTGATTAAC-3′ (SEQ ID NO:47). The PCR fragments were subcloned using the TOPO TA Cloning KIT (Invitrogen) resulting in the plasmids pAAV12-Rep and pAAV12-Cap. Three clones, that were capable of generating recombinant virus, were sequenced with an ABI Prism 3100 Genetic Analyzer (Applied Biosystems, Foster City, Calif.) and FS dye-terminator chemistry (Applied Biosystems).
[0238]Sequence analysis: The sequences of AAV12 rep and cap were compared to sequences in GenBank using BLAST. DNA alignments were performed using the ClustalW multiple sequence alignment tool of the Biology Workbench web based software at http://seqtool.sdsc.edu (SDSC) and MacVector 7 (Accelrys, Burlington, Mass.).
[0239]Generation of recombinant virus: AAV12 (AAVX26) vectors, expressing a nuclear localized green fluorescent protein (GFP), rAAV12GFP, were produced as described above. Briefly, 293T cells were cotransfected with pAAV2-NLS-GFP, pAAV12Rep and pAAV12Cap and the Ad helper plasmids 449B. Recombinant particles were purified by CsC1 gradient centrifugation. DNase-resistant genome copy numbers of the vector stocks were determined by quantitative real-time PCR using the TAQMAN system (Applied Biosystems) with probes specific to the CMV promoter. AAV12-Epo expressing human erythropoietin was generated accordingly by packaging pAAVhEPO with pAAV12Rep and pAAV12Cap.
[0240]Digestion of cell surface sialic acid: Exponentially growing COS cells were plated at a density of 5×103/well in a flat-bottom 96-well plate. Twenty-four hours after seeding, cells were incubated for 30 min with 0.1 and 1 mU of the broad-spectrum neuraminidase from Vibrio cholerae (Calbiochem, La Jolla, Calif.). Cells were then washed with medium and transduced with 1×107particles of rAAV2-GFP, rAAV4-GFP, rAAV5-GFP or rAAV12-GFP. GFP expression, which serves as a surrogate marker for transduction, was detected twenty-four hours later with a fluorescent cell counter (Guava PCA-96, Guava Technologies, Hayward, Calif.).
[0241]Heparin competition assay: COS cells were plated at a density of 5×103/well in a flat-bottom 96-well plate one day prior to transduction. 2×107particles rAAV2-GFP, rAAV12-GFP were preincubated for 1 h at room temperature in medium supplemented with either heparin, mannose or mannosamine (Sigma, St. Louis, Mo.). This pre-incubation mixture was then added and left on the cells for 1 h at 37° C. Cells were then washed with medium and incubated for one day before GFP expression was detected with a fluorescent cell counter (Guava Technologies).
[0242]Protease treatment: COS cells were cultured in a 15-cm diameter culture dish until cells were 80% confluent. Cells were then washed twice with PBS, scraped, resuspended in 10 ml PBS and incubated with 0.05% trypsin (Biosource), or mock (untreated control) for 15 min at 37° C. Cells were then washed twice with medium and seeded at a density of 10,000 cells/well in a 96-well dish. After 1 h culture at 37° C., cells were transduced with 2×107particles rAAV2-GFP, rAAV12-GFP. Transduction efficiency was determined 24 h later by GFP expression detection with a fluorescent cell counter (Guava Technologies).
[0243]Inhibition of glycolipid synthesis: COS cells were plated at a density of 5×103/well in a flat-bottom 96-well plate. Eight hours after seeding, cells were incubated for 40 h with the glucosylceramide synthase inhibitors DL-threo-1-Pheny1-2-palmitoylamino-3-morpholino-1-propanol (PPMP) (Sigma, St. Louis, Mo.). Cells were then washed with medium and transduced with 2×108particles of rAAV2-GFP, rAAV12-GFP and rBAAV-GFP for 1 h. GFP expression was analyzed 48 h after transduction by detection with a fluorescent cell counter (Guava Technologies).
[0244]Neutralization assay: COS cells were seeded at a density of 5×103/well in a 96-well plate 1 day before inoculation. 2×107rAAV2-GFP, rAAV6-GFP and rAAV12-GFP particles that were pre-incubated with serial dilutions of pooled human IgGs (Immune Globuline Intravenous, 10%, Gamunex, BAYER, Leverkusen, Germany) in medium for 1 h at room temperature. Cells were then inoculated with this mixture for 1 h at 37° C. and then washed with medium. Twenty-four hours after transduction, cells were analyzed for GFP expression by flow cytometry (Guava Technologies).
[0245]Animal experiments: Male Balb/c mice were obtained from the Division of Cancer Treatment, NCl, Bethesda, Md. Mice were administered 109particles (suspended in 100 μl of 0.9% NaCl) of either AAV2-hEPO (n=3) or AAV12-hEPO (n=2) by retrograde ductal delivery to both submandibular glands (SG). Two additional groups (n=3 and n=2, respectively) received an equal dose of 109particles (suspended in 100 μl of 0.9% NaCl) in both their tibialis anterior (two injection sites per muscle). A further group (n=3) of naïve mice (administered with 50 μl of 0.9% NaCl to each SG) was included. Mild anesthesia was induced to all participating animals with 1 μl/g body weight of a 60 mg/ml ketamine (Phoenix Scientific, St. Joseph, Mo.) and 8 mg/ml xylazine (Phoenix Scientific) solution given intramuscularly. Blood samples were obtained by orbital bleeding at distinct time points. Hematocrits (Hcts) were determined using micro-hematocrit capillary tubes (Fisher Scientific, Pittsburgh, Pa.). Secretion of hEPO in mouse serum was determined by an ELISA using commercial assay kits (R&D systems, MN, USA). The lower limit of detection was 0.6 mU/ml. Assays were performed according to the manufacturer's instructions.
[0247]Identification of AAV contaminations in ATCC virus isolates: AAV sequences were detected in a Simian Adenovirus 18 strain C676 stock, VR-943, which was isolated from a Vervet monkey (Cercopithecus aethiops). The entire rep and cap coding region of the AAV contamination in VR-943, termed AAV12, has been PCR amplified and subcloned (FIG. 1). Since the rep and cap encoding PCR fragments also contain viral promoter elements, these plasmids could be used as packaging constructs for the generation of recombinant virus. Vectors based on AAV12 were produced by cotransfecting an AAV-2 vector plasmid, encoding a nuclear localized GFP flanked by AAV2 inverted terminal repeats (ITRs), and plasmids encoding AAV12 rep and cap together with an adenovirus helper plasmid. Recombinant viruses were then assayed for transduction activity by inoculating COS cells and assaying for GFP expression by flow cytometry. Three clones, which were capable of generating recombinant virus were sequenced.
[0248]Phylogenetic analysis: The evolutionary relationship among mammalian AAVs and AAV12 was analyzed by ClustalW alignments of genomic, Rep78 and VP1 sequences and plotted as a rooted phylogenetic tree (FIG. 1). The DNA sequence of AAV12 showed highest homology with AAV 11 and AAV4, 83% and 81% respectively, whereas lowest similarity was observed with AAV5 (63%). The Rep78 amino acid sequence of AAV12 demonstrated high homology to AAV13, AAV4, AAV10 and AAV11 with 89% or 88% identity. For the capsid protein VP1, highest homology was observed with AAV11 and AAV4, 84% and 78% respectively, whereas AAV5 VP1 displayed lowest similarity with 53%.
[0249]Heparin competition: Heparan sulfate, a cell surface glycosaminoglycan, which is expressed by virtually all cells, is an attachment receptor for AAV2 (Summerford, C., et al. 1998). AAV2 transduction can be inhibited by heparin, a heparan sulfate analog. Thus, analysis was conducted to determine whether AAV 12 uses heparan sulfate as a receptor by heparin competition experiments. COS cells were transduced with rAAV12 in the presence of increasing amounts of heparin. rAAV2 served as a control. rAAV2 transduction was inhibited to 75% at a concentration of 12.5 μg/ml heparin, whereas no inhibition of rAAV12 was observed at a 80 hold higher heparin concentration of 1000 μg/ml (FIG. 12). Since heparin has no inhibitory potential towards AAV12 transduction, heparan sulfate appears not to be involved in AAV12 transduction.
[0250]Effect of neuraminidase treatment on rAAV12 transduction: Sialic acids, a family of monosaccharides based on N-acetylneuramic acid, are commonly found on the outermost end of glykans and glycolipids. Sialic acids have been identified to serve as receptors for several viruses, including influenza virus, AAV4, AAV5 AAV6 and AAV(VR-355). To analyze, if AAV 12 transduction is sialic acid dependent, the effect of the removal of cell surface sialic acids was studied by neuraminidase treatment on gene transfer (FIG. 134). Enzymatic digestion of COS cells with a broad spectrum neuraminidase from Vibrio cholera inhibited rAAV4 and rAAV5 transduction dose-dependently and blocked gene transfer up to 99% and 97% respectively. AAV12 transduction was unaffected by the enzymatic removal of cell surface sialic acids, indicating that AAV12 does not utilize sialic acid in the transduction process.
[0251]rAAV12 transduction is protease sensitive and does not require glycosphingolipids (GSLs): rBAAV cell entry and transduction depend on gangliosides, glycosphingolipids with sialic acid groups, and are resistant to protease treatment of the cell. To analyze if the rAAV12 receptor is a protein and if glycolipids are involved in the transduction process, the effect of proteolytically digesting cell surface proteins prior to transduction was studied (FIG. 14A). The dependency of rAAV12 transduction on GSLs was studied by incubating cells with DL-threo-1-Phenyl-2-palmitoylamino-3-morpholino-1-propanol (PPMP), a glucosylceramide synthase inhibitors, which act to deplete GSLs from the cell membrane prior to transduction (FIG. 14B). As shown in FIG. 14, AAV12 transduction is protease sensitive, but AAV12 can transduce cells lacking glycosphingolipids.
[0252]rAAV12 is inhibited by extracellular mannosamine: Since rAAV12 transduction does not depend on either heparin or sialic acid, a broad panel of carbohydrates was screened for the ability to interfere with rAAV12 transduction to identify components that might be involved in the rAAV 12 cell interaction. In this assay, mannosamine was identified as compound with inhibitory function against rAAV12. Pre-incubation of rAAV12 with mannosamine prior to transduction resulted in a dose-dependent inhibition of rAAV12 transduction (FIG. 15), while no specific inhibition of rAAV2 was observed. This result indicates that mannosamine could be part of the rAAV12 receptor or attachment factor.
[0253]rAAV12 has a broad tropism in human cancer cell lines: All studied AAVs utilize either sialic acid or heparin as cellular receptor or attachment factor. In contrast, rAAV12 trasduced cells independently of heparin and sialic acid but was inhibited by mannosamine, indicating that rAAV12 interacts in a unique way with the cell. To analyze if unique cell interaction results in a unique tropism, the transduction efficiency of rAAV12 was studied in 13 human cancer cell lines and compared it to rAAV4, a virus that shows 78% homology to rAAV12 in the capsid protein VP1 (FIG. 16). rAAV12 and rAAV4 demonstrated both a broad tropism with overall similar transduction efficiencies, but both viruses had a unique transduction profile. While only rAAV4 transduced HCT10 cells, rAAV12 gene transfer was specific for A549 and HCT15 cells.
[0254]Immunological characterization of rAAV12: Infections with AAV2 are very common and approximately 80% of the human population are seropositive. Neutralization of human AAV serotypes, such as AAV2, AAV3 and AAV5, as well as the simian AAV6 by human serum have been reported. This pre-existing immunity against these AAVs might limit their usability as vectors for gene therapy. To test if rAAV 12 is antigentically distinct from rAAV2 and rAAV6, assays were conducted to determine if IgGs, purified from pooled human serum, contain neutralizing antibodies against the recombinant AAVs and whether there was a difference in the neutralization activity against either rAAV2, AAV6, or rAAV12 (FIG. 17). In this assay the human rAAV2 and simian rAAV6 displayed similar sensitivity to neutralization with the purified pooled IgGs and transduction was inhibited to 50% at a concentration of 0.01 mg/ml IgGs. In contrast, rAAV12 was highly resistant to neutralization by pooled human IgGs, and even at the highest concentration of 0.67mg/ml, a concentration where 100% inhibition of rAAV2 and rAAV6 was observed, rAAV 12 transduction was only reduced by 30%.
[0255]rAAV12 transduces salivary glands and skeletal muscles in vivo: rAAV 12 transduced salivary glands and skeletal muscles with similar efficiency as rAAV2 (FIG. 18).
REFERENCES
[0000]- Allen, J. M., Halbert, C. L. and Miller, A. D.: Improved adeno-associated virus vector production with transfection of a single helper adenovirus gene, E4orf6. Mol Ther 1 (2000) 88-95.
- Amberg, N., A. H. Kidd, K. Edlund, J. Nilsson, P. Pring-Akerblom, and G. Wadell. 2002. Adenovirus type 37 binds to cell surface sialic acid through a charge-dependent interaction. Virology 302:33-43.
- Atchison, R. W., B. C. Casto, and W. M. Hammon. 1965. Adenovirus-Associated Defective Virus Particles. Science 149:754-6.
- Bartlett, J. S., Wilcher, R. and Samulski, R. J.: Infectious entry pathway of adeno-associated virus and adeno-associated virus vectors. J Virol 74 (2000) 2777-85.
- Ben-Israel, H. and Kleinberger, T.: Adenovirus and cell cycle control. Front Biosci 7 (2002) D1369-95.
- Berns, K. I. 1996. Parvoviridae: the viruses and their replication, p. 2173-2197. In F. B. N., K. D. M., and H. P. M. (ed.), Fields virology, 3rd ed. Lippincott-Raven Publishers, Philadelphia, Pa.
- Blacklow, N. R., Hoggan, M. D. and Rowe, W. P.: Isolation of adenovirus-associated viruses from man. Proc Natl Acad Sci USA 58 (1967) 1410-5.
- Blacklow, N. R., Hoggan, M. D. and Rowe, W. P.: Serologic evidence for human infection with adenovirus-associated viruses. J Natl Cancer Inst 40 (1968) 319-27.
- Bossis, I. and Chiorini, J. A.: Cloning of an Avian Adeno-Associated Virus (AAAV) and Generation of Recombinant AAAV Particles. J Virol 77 (2003) 6799-810.
- Carter, B. J., B. A. Antoni, and D. F. Klessig. 1992. Adenovirus containing a deletion of the early region 2A gene allows growth of adeno-associated virus with decreased efficiency. Virology 191:473-6.
- Carter, B. J., C. A. Laughlin, L. M. de la Maza, and M. Myers. 1979. Adenoassociated virus autointerference. Virology 92:449-62.
- Casto, B. C., J. A. Armstrong, R. W. Atchison, and W. M. Hammon. 1967. Studies on the relationship between adeno-associated virus type 1 (AAV-1) and adenoviruses. II. Inhibition of adenovirus plaques by AAV; its nature and specificity. Virology 33:452-8.
- Casto, B. C., R. W. Atchison, and W. M. Hammon. 1967. Studies on the relationship between adeno-associated virus type I (AAV-1) and adenoviruses. I. Replication of AAV-1 in certain cell cultures and its effect on helper adenovirus. Virology 32:52-9.
- Chang, L. S, and Shenk, T.: The adenovirus DNA-binding protein stimulates the rate of transcription directed by adenovirus and adeno-associated virus promoters. J Virol 64 (1990) 2103-9.
- Chiorini, J. A., F. Kim, et al. (1999). “Cloning and characterization of adeno-associated virus type 5.” J Virol 73(2): 1309-19.
- Chiorini, J. A., L. Yang, Y. Liu, B. Safer, and R. M. Kotin. 1997. Cloning of adeno-associated virus type 4 (AAV4) and generation of recombinant AAV4 particles. J Virol 71:6823-33.
- Chiorini, J. A., Wiener, S. M., Owens, R. A., Kyostio, S. R., Kotin, R. M. and Safer, B.: Sequence requirements for stable binding and function of Rep68 on the adeno-associated virus type 2 inverted terminal repeats. J Virol 68 (1994) 7448-57.
- Davidson, B. L., C. S. Stein, et al. (2000). “Recombinant adeno-associated virus type 2, 4, and 5 vectors: transduction of variant cell types and regions in the mammalian central nervous system.” Proc Natl Acad Sci USA 97(7): 3428-32.
- Derby, M. L., M. Sena-Esteves, et al. (1999). “Gene transfer into the mammalian inner ear using HSV-1 and vaccinia virus vectors.” Hear Res 134(1-2): 1-8.
- Di Pasquale, G., and J. A. Chiorini. 2003. PKA/PrKX activity is a modulator of AAV/adenovirus interaction. EMBO J. 22:1716-24.
- Di Pasquale, G., B. L. Davidson, et al. (2003). “Identification of PDGFR as a receptor for AAV5 transduction.” Nat Med 9(10): 1306-12.
- Di Pasquale, G., Rzadzinska, A., Schneider, M. E., Bossis, I., Chiorini, J. A., Kachar, B., A novel bovine virus efficiently transduces inner ear neuroepithelial cells, Manuscript submitted.
- Erles, K., Sebokova, P. and Schlehofer, J. R.: Update on the prevalence of serum antibodies (IgG and IgM) to adeno-associated virus (AAV). J Med Virol 59 (1999) 406-11.
- Gao, G., L. H. Vandenberghe, M. R. Alvira, Y. Lu, R. Calcedo, X. Zhou, and J. M. Wilson.
- 2004. Clades of Adeno-associated viruses are widely disseminated in human tissues. J Virol 78:6381-8.
- Gao, G. P., Alvira, M. R., Wang, L., Calcedo, R., Johnston, J. and Wilson, J. M.: Novel adeno-associated viruses from rhesus monkeys as vectors for human gene therapy. Proc Natl Acad Sci USA 99 (2002) 11854-9.
- Georg-Fries, B., S. Biederlack, J. Wolf, and H. zur Hausen. 1984. Analysis of proteins, helper dependence, and seroepidemiology of a new human parvovirus. Virology 134:64-71.
- Grimm, D. and M. A. Kay (2003). “From virus evolution to vector revolution: use of naturally occurring serotypes of adeno-associated virus (AAV) as novel vectors for human gene therapy.” Curr Gene Ther 3(4): 281-304.
- Halbert, C. L., J. M. Allen, and A. D. Miller. 2001. Adeno-associated virus type 6 (AAV6) vectors mediate efficient transduction of airway epithelial cells in mouse lungs compared to that of AAV2 vectors. J Virol 75:6615-24.
- He, D. Z., J. Zheng, et al. (2001). “Development of acetylcholine receptors in cultured outer hair cells.” Hear Res 162(1-2): 113-25.
- Heister, T., Heid, I. Ackermann, M., Fraefel, C. Herpes simplex virus type 1/adeno-associated virus hybrid vectors mediate site-specific integration at the adeno-associated virus preintegration site, AAVS1, on human chromosome 19. J. Virol. 2002 July; 76(14):7163-73.
- Hoggan, M. D. 1970. Adenovirus associated viruses. Prog Med Virol 12:211-39.
- Hoggan, M. D., N. R. Blacklow, and W. P. Rowe. 1966. Studies of small DNA viruses found in various adenovirus preparations: physical, biological, and immunological characteristics. Proc Natl Acad Sci USA 55:1467-74.
- Holt, J. R. (2002). “Viral-mediated gene transfer to study the molecular physiology of the Mammalian inner ear.” Audiol Neurootol 7(3): 157-60.
- Holt, J. R., D. C. Johns, et al. (1999). “Functional expression of exogenous proteins in mammalian sensory hair cells infected with adenoviral vectors.” J Neurophysiol 81(4): 1881-8.
- Hull, R. N., and J. R. Minner. 1957. New viral agents recovered from tissue cultures of monkey kidney cells. II. Problems of isolation, and identification. Ann N Y Acad Sci 67:413-23.
- Hull, R. N., J. R. Minner, and C. C. Mascoli. 1958. New viral agents recovered from tissue cultures of monkey kidney cells. III. Recovery of additional agents both from cultures of monkey tissues and directly from tissues and excreta. Am J Hyg 68:31-44.
- Hull, R. N., J. R. Minner, and J. W. Smith. 1956. New viral agents recovered from tissue cultures of monkey kidney cells. I. Origin and properties of cytopathogenic agents S.V.1, S.V.2, S.V.4, S.V.5, S.V.6, S.V.11, S.V.12 and S.V.15. Am J Hyg 63:204-15.
- Janik, J. E., Huston, M. M., Cho, K. and Rose, J. A.: Efficient synthesis of adeno-associated virus structural proteins requires both adenovirus DNA binding protein and VA I RNA. Virology 168 (1989) 320-9.
- Jero J, Mhatre A N, Tseng C J, Stem R E, Coling D E, Goldstein J A, Hong K, Zheng W W, Hogue A T, Lalwani A K. Cochlear gene delivery through an intact round window membrane in mouse. Hum Gene Ther. 2001 Mar. 20; 12(5):539-48.
- Kaludov, N., K. E. Brown, et al. (2001). “Adeno-associated virus serotype 4 (AAV4) and AAV5 both require sialic acid binding for hemagglutination and efficient transduction but differ in sialic acid linkage specificity.” J Virol 75(15): 6884-93.
- Kanzaki, S., K. Ogawa, et al. (2002). “Transgene expression in neonatal mouse inner ear explants mediated by first and advanced generation adenovirus vectors.” Hear Res 169(1-2): 112-20.
- Katano H, Afione S, Schmidt M, Chiorini J A. Identification of adeno-associated virus contamination in cell and virus stocks by PCR. Biotechniques. 2004 April; 36(4):676-80.
- Kelsell, D. P., Dunlop, J., Stevens, H. P., Lench, N. J., Liang, J. N., Parry, G., Mueller, R. F., Leigh, I. M. Connexin 26 mutations in hereditary non-syndromic sensorineural deafness. Nature. 1997 May 1; 387(6628):80-3
- Kern, A., K. Schmidt, C. Leder, O. J. Muller, C. E. Wobus, K. Bettinger, C. W. Von der Lieth, J. A. King, and J. A. Kleinschmidt. 2003. Identification of a heparin-binding motif on adeno-associated virus type 2 capsids. J Virol 77:11072-81.
- Li Duan, M., T. Bordet, et al. (2002). “Adenoviral and adeno-associated viral vector mediated gene transfer in the guinea pig cochlea.” Neuroreport 13(10): 1295-9.
- Luchsinger, E., Strobbe, R., Dekegel, D. and Wellemans, G.: Use of B-IV zonal rotor centrifugation as a simple tool for the separation of adeno-associated X 7 virus (AAVX 7) from helper adenoviruses. Arch Gesamte Virusforsch 33 (1971) 251-8.
- Luchsinger, E., Strobbe, R., Wellemans, G., Dekegel, D. and Sprecher-Goldberger, S.: Haemagglutinating adeno-associated virus (AAV) in association with bovine adenovirus type 1. Brief report. Arch Gesamte Virusforsch 31 (1970) 390-2.
- Luebke, A. E., J. D. Steiger, et al. (2001). “A modified adenovirus can transfect cochlear hair cells in vivo without compromising cochlear function.” Gene Ther 8(10): 789-94.
- Luebke, A. E., P. K. Foster, et al. (2001). “Cochlear function and transgene expression in the guinea pig cochlea, using adenovirus- and adeno-associated virus-directed gene transfer.” Hum Gene Ther 12(7): 773-81.
- McPherson, R. A., L. J. Rosenthal, and J. A. Rose. 1985. Human cytomegalovirus completely helps adeno-associated virus replication. Virology 147:217-22.
- Meyers, C., M. Mane, N. Kokorina, S. Alam, and P. L. Hermonat. 2000. Ubiquitous human adeno-associated virus type 2 autonomously replicates in differentiating keratinocytes of a normal skin model. Virology 272:338-46.
- Mori, S., L. Wang, T. Takeuchi, and T. Kanda. 2004. Two novel adenoassociated viruses from cynomolgus monkey: pseudotyping characterization of capsid protein. Virology 330:375-83.
- Mouw, M. B. and Pintel, D. J.: Adeno-associated virus RNAs appear in a temporal order and their splicing is stimulated during coinfection with adenovirus. J Virol 74 (2000) 9878-88.
- Muramatsu, S., H. Mizukami, et al. (1996). “Nucleotide sequencing and generation of an infectious clone of adeno-associated virus 3.” Virology 221(1): 208-17.
- Myrup, A. C., Mohanty, S. B. and Hetrick, F. M.: Isolation and characterization of adeno-associated viruses from bovine adenovirus types 1 and 2. Am J Vet Res 37 (1976) 907-10.
- Naz, S., Griffith, A. J., Riazuddin, S., Hampton, L. L., Battey, J. F. Jr, Khan, S. N., Riazuddin, S., Wilcox, E. R., Friedman, T. B. Mutations of ESPN cause autosomal recessive deafness and vestibular dysfunction. J Med. Genet. 2004 August,41(8):591-5.
- No D, Yao T P, Evans R M. (1996). Ecdysone-inducible gene expression in mammalian cells and transgenic mice. Proc Natl Acad Sci USA. 93(8):3346-51.
- Ogston, P., K. Raj, and P. Beard. 2000. Productive replication of adenoassociated virus can occur in human papillomavirus type 16 (HPV-16) episomecontaining keratinocytes and is augmented by the HPV-16 E2 protein. J Virol 74:3494-504.
- Opie, S. R., K. H. Warrington, Jr., et al. (2003). “Identification of amino acid residues in the capsid proteins of adeno-associated virus type 2 that contribute to heparan sulfate proteoglycan binding.” J Virol 77(12): 6995-7006.
- Parks, W. P., J. L. Melnick, R. Rongey, and H. D. Mayor. 1967. Physical assay and growth cycle studies of a defective adeno-satellite virus. J Virol 1:171-80.
- Rabinowitz J E, Bowles D E, Faust S M, Ledford J G, Cunningham S E, Samulski R J. Cross-dressing the virion: the transcapsidation of adeno-associated virus serotypes functionally defines subgroups. J. Virol. 2004 May; 78(9):4421-32.
- Reddy, V. S., P. Natarajan, B. Okerberg, K. Li, K. V. Damodaran, R. T. Morton, C. L. Brooks, 3rd, and J. E. Johnson. 2001. Virus Particle Explorer (VIPER), a website for virus capsid structures and their computational analyses. J Virol 75:11943-7.
- Richardson, W. D., and H. Westphal. 1984. Requirement for either early region 1a or early region 1b adenovirus gene products in the helper effect for adenoassociated virus. J Virol 51:404-10.
- Rutledge, E. A., C. L. Halbert, and D. W. Russell. 1998. Infectious clones and vectors derived from adeno-associated virus (AAV) serotypes other than AAV type 2. J Virol 72:309-19.
- Rzadzinska, A. K., M. E. Schneider, et al. (2004). “An actin molecular treadmill and myosins maintain stereocilia functional architecture and self-renewal.” J Cell Biol 164(6): 887-97.
- Saffer, L. D., R. Gu, et al. (1996). “An RT-PCR analysis of mRNA for growth factor receptors in damaged and control sensory epithelia of rat utricles.” Hear Res 94(1-2): 14-23.
- Samulski, R. J., and T. Shenk. 1988. Adenovirus E1B 55-Mr polypeptide facilitates timely cytoplasmic accumulation of adeno-associated virus mRNAs. J Virol 62:206-10.
- Samulski, R. J., K. I. Berns, M. Tan, and N. Muzyczka. 1982. Cloning of adeno-associated virus into pBR322: rescue of intact virus from the recombinant plasmid in human cells. Proc Natl Acad Sci USA 79:2077-81.
- Sanlioglu, S., Benson, P. K., Yang, J., Atkinson, E. M., Reynolds, T. and Engelhardt, J. F.: Endocytosis and nuclear trafficking of adeno-associated virus type 2 are controlled by rac1 and phosphatidylinositol-3 kinase activation. J Virol 74 (2000) 9184-96.
- Schlehofer, J. R., M. Ehrbar, and H. zur Hausen. 1986. Vaccinia virus, herpes simplex virus, and carcinogens induce DNA amplification in a human cell line and support replication of a helpervirus dependent parvovirus. Virology 152:110-7.
- Schmidt, M., H. Katano, et al. (2004). “Cloning and characterization of a bovine adeno-associated virus.” J Virol 78(12): 6509-16.
- Schmidt, M., H. Katano, I. Bossis, and J. A. Chiorini. 2004. Cloning and characterization of a bovine adeno-associated virus. J Virol 78:6509-16.
- Schneider, M. E., I. A. Belyantseva, et al. (2002). “Rapid renewal of auditory hair bundles.” Nature 418(6900): 837-8.
- Schwede, T., J. Kopp, N. Guex, and M. C. Peitsch. 2003. SWISS-MODEL: An automated protein homology-modeling server. Nucleic Acids Res 31:3381-5.
- Seiler, M. P., C. L. Halbert, J. A. Chiorini, A. D. Miller, and J. Zabner. 2002. AAV5 and AAV6 Mediate Gene Transfer to Human Airway Epthelia Via Different Receptors. Mol. Ther. 5:540.
- Shou, J., J. L. Zheng, et al. (2003). “Robust generation of new hair cells in the mature mammalian inner ear by adenoviral expression of Hathl.” Mol Cell Neurosci 23(2): 169-79.
- Smith, R. H., and R. M. Kotin. 2002. Adeno-associated virus as a transposable element. In R. C. Nancy L. Craig, Martin Gellert, and Alan M. Lambowitz (ed.), Mobile DNA II. ASM Press.
- Smith, R. H., S. A. Afione, and R. M. Kotin. 2002. Transposase-mediated construction of an integrated adeno-associated virus type 5 helper plasmid. Biotechniques 33:204-6, 208, 210-1.
- Sobkowicz, H. M., J. M. Loftus, et al. (1993). “Tissue culture of the organ of Corti.” Acta Otolaryngol Suppl 502: 3-36.
- Srivastava, A., E. W. Lusby, and K. I. Berns. 1983. Nucleotide sequence and organization of the adeno-associated virus 2 genome. J Virol 45:555-64.
- Staecker H, Li D, O'Malley B W Jr, Van De Water T R. Gene expression in the mammalian cochlea: a study of multiple vector systems. Acta Otolaryngol. 2001 January; 121(2):157-63.
- Stracker, T. H., G. D. Cassell, P. Ward, Y. M. Loo, B. van Breukelen, S. D. Carrington-Lawrence, R. K. Hamatake, P. C. van der Vliet, S. K. Weller, T. Melendy, and M. D. Weitzman. 2004. The Rep protein of adeno-associated virus type 2 interacts with single-stranded DNA-binding proteins that enhance viral replication. J Virol 78:441-53.
- Summerford, C. and R. J. Samulski (1998). “Membrane-associated heparan sulfate proteoglycan is a receptor for adeno-associated virus type 2 virions.” J Virol 72(2): 1438-45.
- Superti, F., M. L. Marziano, A. Tinari, and G. Donelli. 1993. Effect of polyions on the infectivity of SA-11 rotavirus in LCC-MK2 cells. Comp Immunol Microbiol Infect Dis 16:55-62.
- Suzuki, H., Y. Katori, et al. (1995). “Carbohydrate distribution in the living utricular macula of the guinea pig detected by lectins.” Hear Res 87(1-2): 32-40.
- Tratschin, J. D., M. H. West, T. Sandbank, and B. J. Carter. 1984. A human parvovirus, adeno-associated virus, as a eucaryotic vector: transient expression and encapsidation of the procaryotic gene for chloramphenicol acetyltransferase. Mol Cell Biol 4:2072-81.
- Walters, R. W., S. M. Yi, S. Keshavjee, K. E. Brown, M. J. Welsh, J. A. Chiorini, and J. Zabner. 2001. Binding of adeno-associated virus type 5 to 2,3-linked sialic acid is required for gene transfer. J Biol Chem 276:20610-6.
- Walz, C., A. Deprez, T. Dupressoir, M. Durst, M. Rabreau, and J. R. Schlehofer. 1997. Interaction of human papillomavirus type 16 and adenoassociated virus type 2 co-infecting human cervical epithelium. J Gen Virol 78 (Pt 6):1441-52.
- Wang, X. S., and A. Srivastava. 1998. Rescue and autonomous replication of adeno-associated virus type 2 genomes containing Rep-binding site mutations in the viral p5 promoter. J Virol 72:4811-8.
- Ward, P., F. B. Dean, M. E. O'Donnell, and K. I. Berns. 1998. Role of the adenovirus DNA-binding protein in in vitro adeno-associated virus DNA replication. J Virol 72:420-7.
- Weindler, F. W., and R. Heilbronn. 1991. A subset of herpes simplex virus replication genes provides helper functions for productive adeno-associated virus replication. J Virol 65:2476-83.
- Xiao, W., N. Chirmule, S. C. Berta, B. McCullough, G. Gao, and J. M. Wilson. 1999. Gene therapy vectors based on adeno-associated virus type 1. J Virol 73:3994-4003.
- Yakinoglu, A. 0., R. Heilbronn, A. Burkle, J. R. Schlehofer, and H. zur Hausen. 1988. DNA amplification of adeno-associated virus as a response to cellular genotoxic stress. Cancer Res 48:3123-9.
- Yakobson, B., T. A. Hrynko, M. J. Peak, and E. Winocour. 1989. Replication of adeno-associated virus in cells irradiated with UV light at 254 nm. J Virol 63:1023-30.
- Yamano, S., Huang, L. Y., Ding, C., Chiorini, J. A., Goldsmith, C. M., Wellner, R. B., Golding, B., Kotin, R. M., Scott, D. E. and Baum, B. J.: Recombinant adeno-associated virus serotype 2 vectors mediate stable interleukin 10 secretion from salivary glands into the bloodstream. Hum Gene Ther 13 (2002) 287-98.
- Zabner, J., Seiler, M., Walters, R., Kotin, R. M., Fulgeras, W., Davidson, B. L., Chiorini, J. A. Adeno-associated virus type 5 (AAV5) but not AAV2 binds to the apical surfaces of airway epithelia and facilitates gene transfer. J. Virol. 2000 April; 74(8):3852-8.