[1]Группа изобретений относится к области генной инженерии, молекулярной и клеточной биологии и предназначена для создания универсальной матрицы в виде плазмидной ДНК для получения искусственных матричных РНК (мРНК), обеспечивающих высокий уровень экспрессии закодированных целевых генов в клетках и тканях человека и животных.
[2]Технологии мРНК в настоящее время активно применяются для разработки новых вариантов профилактических противовирусных вакцин, новых терапевтических средств, персонализированных и «универсальных» противораковых мРНК-вакцин. Ключевыми ограничениями использования и внедрения этой технологии являются низкая стабильность мРНК и возможный низкий уровень синтеза целевого белка. На решение указанных ограничений направлено представленное изобретение.
[3]Включение в состав мРНК нетранслируемых областей (UTRs, от англ, untranslated regions), фланкирующих открытую рамку считывания с 5'- и 3'- концов, позволяет значительно усилить уровень трансляции этих мРНК в клетках млекопитающих. Наиболее часто используемыми вариантами парных последовательностей UTR, которые были использованы при конструировании искусственных мРНК и описаны в научной литературе, являются UTR β-глобина [1] (гена НВВ человека), UTR α-глобина (гена НВА1 человека) [1], UTR гемоглобина α [2].
[4]Включение последовательностей 5'-UTR усиливает трансляцию, не только защищая кодирующие последовательности от нуклеаз, но и привлекая рибосомальный аппарат к сайту начала трансляции (TSS, от англ, translation start site). Был проведен отбор последовательностей 5'-UTR («5'-UTR-4») с высокой вероятностью посадки рибосомы с использованием методов машинного обучения [3]. Использование последовательностей, выбранных в исследовании с использованием обучающихся нейросетей, позволило повысить уровень трансляции.
[5]3'-UTR обеспечивают защиту от нуклеаз, включая предотвращение деградации поли(А)-хвоста, которая может влиять на стабильность мРНК, на период полураспада мРНК и выход белка в результате синтеза на рибосомах. Одним из часто используемых вариантов UTRs являются 5'- и 3'-UTR β-глобина (гена НВВ человека), для которых была продемонстрирована эффективность в ряде работ как для мРНК репортерных генов, так и для других моделей, например, вирусных антигенов [1].
[6]Ранее было показано, что включение химерной 3'-UTR AES-mtRNR1, которая состоит из области 16S рибосомальной рРНК архей (AES - Amino-terminal Enhancer of Split) и части митохондриальной рРНК (mtRNR1) среди всех проанализированных в исследовании вариантов структур мРНК позволяет в 3 раза повысить уровень трансляции in vitro по сравнению со стандартными нетранслируемыми областями β-глобина [1].
[7]Ранее не было предложено структур мРНК, содержащих в своем составе обе вышеназванные нетранслируемые области, а также конструкций, позволяющих производить синтез данных мРНК.
[8]Задачей изобретения является получение плазмидных ДНК, которые являются матрицами для ферментативного синтеза мРНК, содержащих в своем составе нетранслируемые области 5'-UTR-4 и 3'-UTR AES-mtRNR1, обеспечивающие высокий уровень синтеза белка в клетках человека и животных.
[9]Технический результат: повышение эффективности трансляции искусственных матричных РНК, а также расширение ассортимента рекомбинантных плазмид, кодирующих синтез искусственных матричных РНК, обеспечивающих высокий уровень экспрессии целевых закодированных генов в клетках и тканях человека и животных.
[10]Указанный результат достигается включением в плазмиду pCMV6-Entry [4], графическая карта которой представлена в источнике [5], следующих фрагментов: промотора РНК-полимеразы фага Т7 с тринуклеотидом для встройки кэп-аналога и со вспомогательной последовательностью, 5'-нетранслируемой области 5'-UTR-4, полилинкера, содержащего сайты рестрикции для ряда рестриктаз (EcoRI, SalI, Sfr274I, CciNI, PspLI, PspOMI, ApaI, HindIII, BamHI), гибридной 3'-нетранслируемой области 3'-UTR-AES-mtRNR1 и сайта рестрикции XbaI.
[11]Сущность заявляемого изобретения заключается в следующем.
[12]Предложены плазмидные генетические конструкции pCMV6_T7_synthUTR_AGN/GNN, содержащие в своем составе последовательности нетранслируемых областей 5'-UTR-4 и 3'-UTR-AES-mtRNR1, фланкирующие полилинкер, содержащий сайты рестрикции для ряда рестриктаз: EcoRI, SalI, Sfr274I, CciNI, PspLI, PspOMI, ApaI, HindIII, BamHI. Для применения представленных плазмид осуществляется стадия рестрикции и клонирования для включения последовательности требуемой кодирующей открытой рамки считывания, наработка полученных плазмид в бактериальных клетках, выделение и очистка для последующей линеаризации методом рестрикции и применения в качестве матриц в транскрипции in vitro с использованием РНК-полимеразы фага Т7.
[13]Исходным генетическим материалом для конструирования плазмид pCMV6_T7_synthUTR_AGN/GNN является плазмида pBLSK_synth_AGG, которая была получена следующим образом: в плазмиду pBluescript SK (+) по сайтам AbsI и BstXI (СибЭнзайм, Россия) были встроены последовательности промотора РНК полимеразы фага Т7 с нуклеотидами для встройки кэп-аналога и вспомогательной последовательностью, 5'-UTR-4, MCS и 3'-UTR-AES-mtRNR1. Последовательности промотора РНК полимеразы фага Т7 с нуклеотидами для встройки кэп-аналога и вспомогательной последовательностью были выбраны из группы:
[14]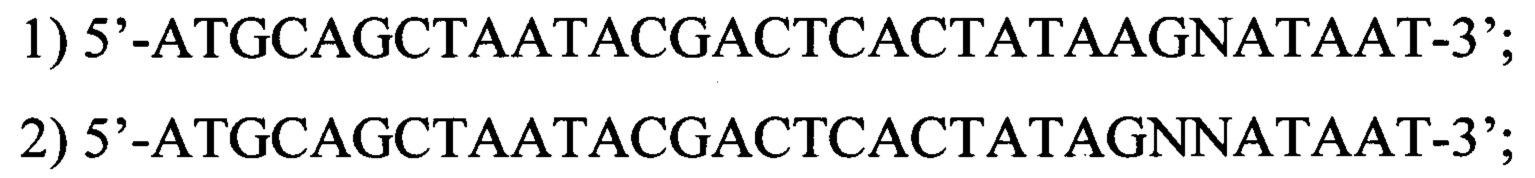
[15]где N - любой нуклеотид.
[16]Вспомогательные последовательности были выбраны в из источника [6]. Последовательности высокоэффективных нетранслируемых областей 5'-UTR-4 и 3'-UTR-AES-mtRNR1 были выбраны из источников [3] и [7], соответственно. Сразу после последовательности 3'-UTR-AES-mtRNR1 был добавлен сайт рестрикции фермента XbaI (СибЭнзайм, Россия) для линеаризации ДНК-матрицы перед синтезом мРНК.
[17]Последовательность MCS (от англ. Multiple Cloning Site) была спроектирована с использованием сайтов рестрикции нескольких наиболее доступных рестриктаз (EcoRI, SalI, Sfr274I, CciNI, PspLI, PspOMI, ApaI, HindIII, BamHI) (СибЭнзайм, Россия).
[18]Далее для получения плазмид pCMV6_T7_synthUTR_AGN/GNN было произведено клонирование последовательностей промотора РНК-полимеразы фага Т7, 5'-UTR-4, MCS и 3'-UTR-AES-mtRNR1 из плазмиды pBLSK synth AGG в плазмиду pCMV6-Entry по сайтам Psp124BI и XmaI (СибЭнзайм, Россия). При этом область, кодирующая целевой белок, может представлять собой последовательность, кодирующую вакцинный антиген, антитело, терапевтический белок или фермент.
[19]Изобретение иллюстрируется следующими фигурами:
[20]Фиг. 1. Графическая карта оригинальной рекомбинантной плазмидной ДНК pCMV6_T7_synthUTR_AGN, содержащей в своем составе нетранслируемые области 5'-UTR-4 и 3'-UTR AES-mtRNR1.
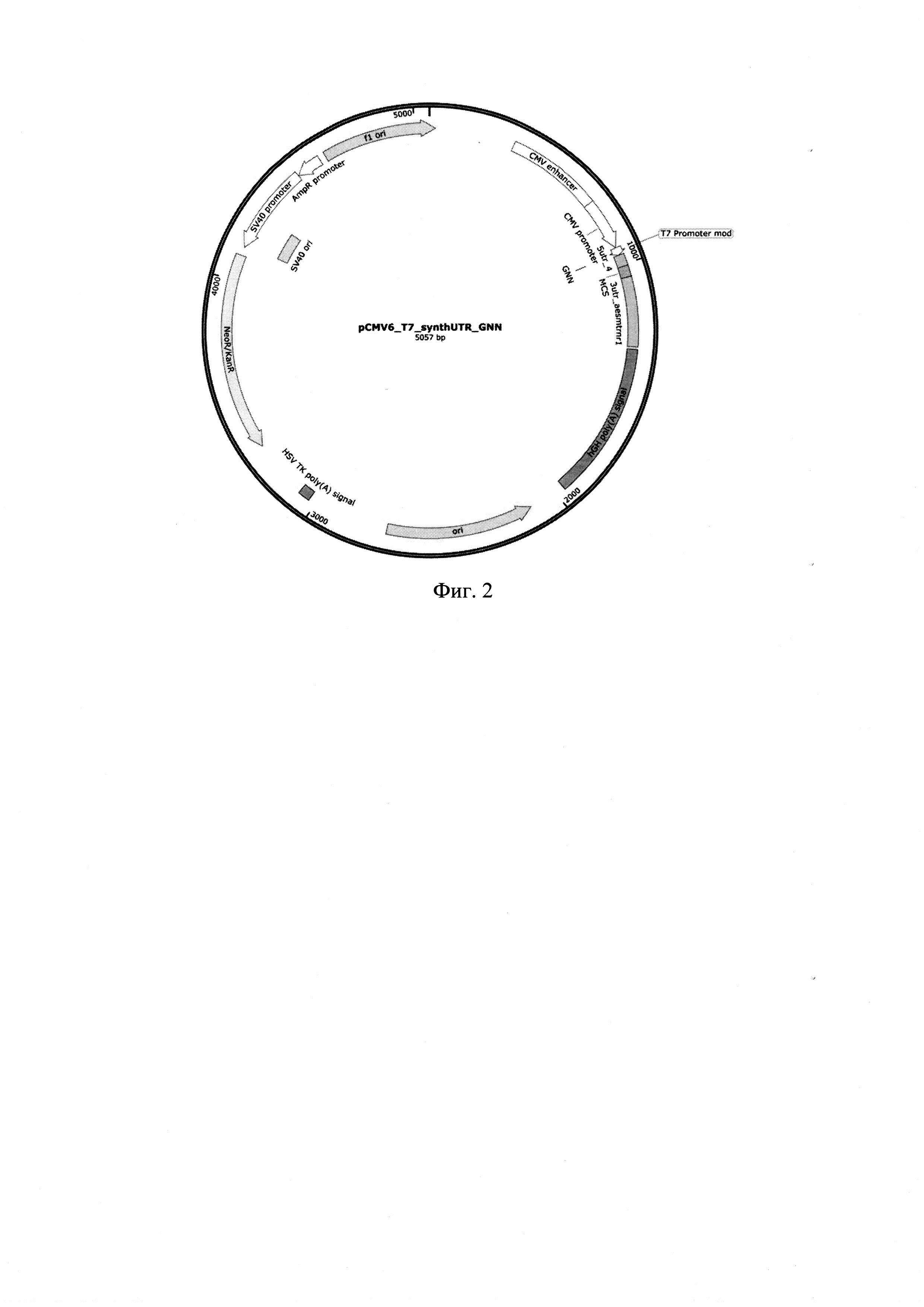
[21]Фиг. 2. Графическая карта оригинальной рекомбинантной плазмидной ДНК pCMV6_T7_synthUTR_GNN, содержащей в своем составе нетранслируемые области 5'-UTR-4 и 3'-UTR AES-mtRNR1.
[22]Фиг.3. Получение мРНК Luc2 и hMGFP с помощью in vitro транскрипции по матрице линеаризованных плазмид pCMV6_T7_synthUTR_AGN_Luc2, pCMV6_T7_synthUTR_AGN_hMGFP, pCMV6_T7_synthUTR_GNN_Luc2, pCMV6_T7_synthUTR_GNN_hMGFP и проверка их качества с помощью электрофореза в 1.5%-ном агарозном геле.
[23]Фиг. 4. Анализ люминесцентного сигнала лизатов клеток НЕК293Т/17, трансфицированных мРНК Luc2, кэпированными кэп-аналогами ARCA или AG, с нетранслируемыми областями β-глобина, либо содержащими в своем составе комбинацию 5'-UTR-4 и 3'-UTR AES-mtRNR1, с помощью флюоресцентного ридера ClarioStar.
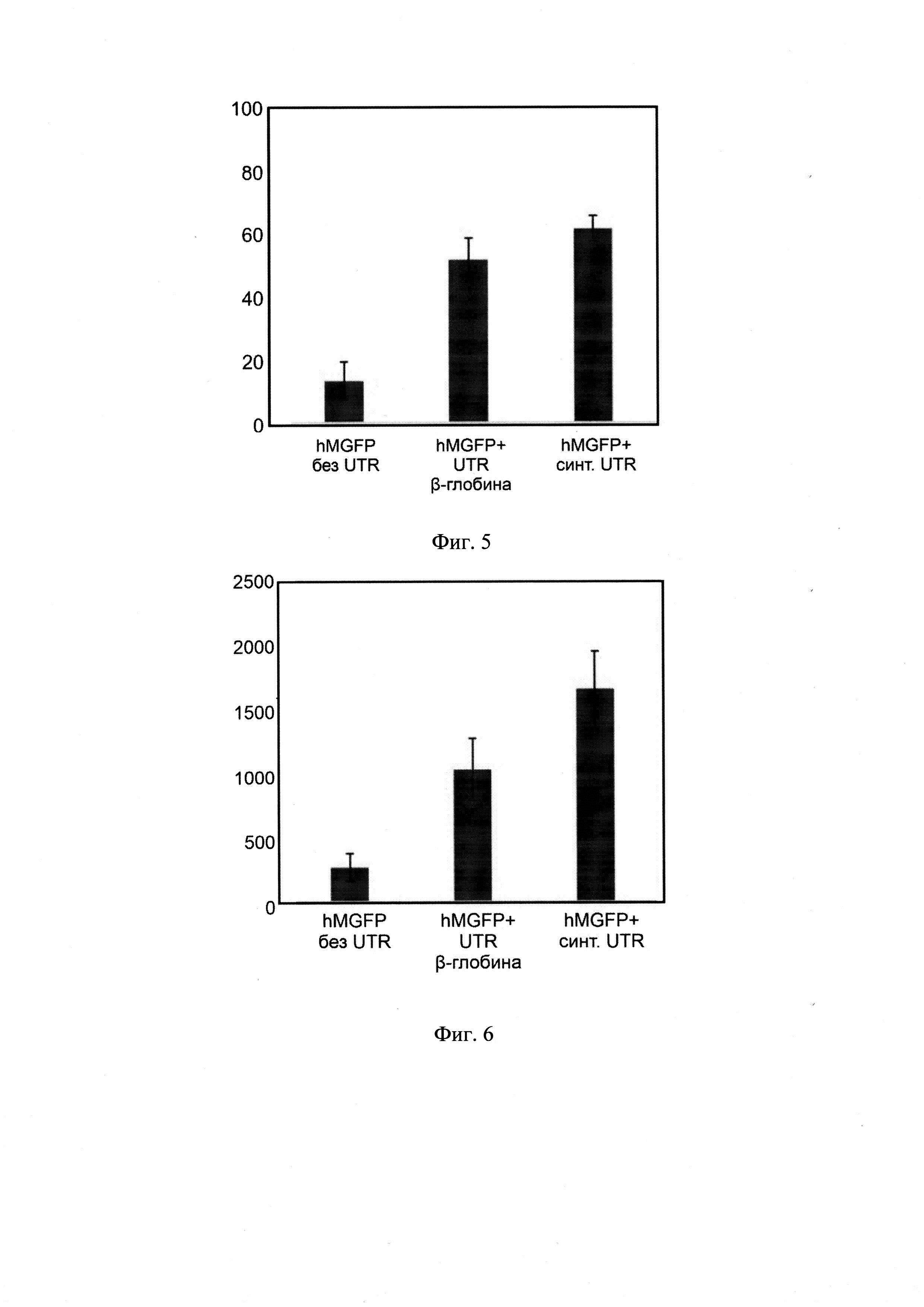
[24]Фиг. 5. Анализ количества GFP+ клеток НЕК293Т/17, трансфицированных мРНК hMGFP, кэпированными кэп-аналогом AG, с нетранслируемыми областями β-глобина, либо содержащими в своем составе комбинацию 5'-UTR-4 и 3'-UTR AES-mtRNR1, с помощью проточной цитометрии на проточном цитометре BD FACSCanto II.
[25]Фиг. 6. Анализ относительного уровня флюоресценции клеток НЕК293Т/17, трансфицированных мРНК hMGFP, кэпированными кэп-аналогом AG, с нетранслируемыми областями β-глобина, либо содержащими в своем составе комбинацию 5'-UTR-4 и 3'-UTR AES-mtRNR1, с помощью проточной цитометрии на проточном цитометре BD FACSCanto II.
[26]Фиг. 7. Анализ люминесцентного сигнала in vivo препаратов мРНК Luc2, содержащих в своем составе кэп AG, с нетранслируемыми областями β-глобина, либо с комбинацией 5'-UTR-4 и 3'-UTR AES-mtRNR1, с помощью прижизненной детекции в аппарате IVIS Lumina Х5.
[27]Для лучшего понимания сущности предлагаемого изобретения, оно иллюстрируется следующими примерами конкретного осуществления.
[28]Пример 1. Получение рекомбинантной плазмидной ДНК pCMV6_T7_synthUTR_AGN, содержащей в своем составе нетранслируемые области 5'-UTR-4 и 3'-UTR AES-mtRNR1.
[29]Исходным генетическим материалом для конструирования плазмиды pCMV6_T7_synthUTR_AGN (фиг.1) является плазмида pBLSK synth AGG, которая была получена следующим образом: в плазмиду pBluescript SK (+) по сайтам AbsI и BstXI (СибЭнзайм, Россия) была встроена последовательность промотора РНК полимеразы фага Т7 с нуклеотидами для встройки кэп-аналога (5'-AGN-3', где N - любой нуклеотид) и вспомогательной последовательностью 5'-АТААТ-3', 5'-UTR-4, MCS и 3'-UTR-AES-mtRNR1.
[30]Последовательности высокоэффективных нетранслируемых областей 5'-UTR-4 и 3'-UTR-AES-mtRNR1 были выбраны из источников [3] и [7], соответственно. Сразу после последовательности 3'-UTR-AES-mtRNR1 был добавлен сайт рестрикции фермента XbaI (СибЭнзайм, Россия) для линеаризации ДНК-матрицы перед синтезом мРНК.
[31]Последовательность MCS (от англ. Multiple Cloning Site) была спроектирована с использованием сайтов рестрикции нескольких наиболее доступных рестриктаз (EcoRI, Sail, Sfr274I, CciNI, PspLI, PspOMI, ApaI, HindIII, BamHI) (СибЭнзайм, Россия).
[32]Далее для получения плазмиды pCMV6_T7_synthUTR_AGN было произведено клонирование последовательностей промотора РНК-полимеразы фага Т7, 5'-UTR-4, MCS и 3'-UTR-AES-mtRNR1 из плазмиды pBLSK synth AGN в плазмиду pCMV6-Entry по сайтам Psp124BI и XmaI (СибЭнзайм, Россия). Для этого готовили отдельные реакционные смеси объемом 20 мкл, содержащие 4 мкг плазмидной ДНК (pBLSK_synth_AGN или pCMV6-Entry), 2 мкл 10х FastDigest Buffer (Thermofisher, В64), 160 е.а. Psp124BI (SibEnzyme, SE-E107), 12 е.а. XmaI (SibEnzyme, SE-E233), вода DEPC до конечного объема. Смеси инкубировали в течение ночи при 37°С в термостате и затем инактивировали при 65°С в течение 20 минут.
[33]Инактивированные реакционные смеси разделяли в препаративном 0,8% агарозном геле с добавлением бромистого этидия. Целевые фрагменты вектора (4645 п. н.) и вставки (412 п. н.) были выделены с использованием набора N-Gel (Биолабмикс, Россия).
[34]Далее была проведена реакция лигирования по следующему протоколу: смешали реакционную смесь объемом 20 мкл (ДНК вектора и вставки в молярном соотношении 1 к 5, 2 мкл 10х Т4 DNA Ligase Buffer (SibEnzyme, В302), 200 Е.А. Т4 DNA Ligase (SibEnzyme, Россия), вода DEPC до конечного объема), инкубировали в течение ночи при 4°С, затем инактивировали при 65°С в течение 10 минут и охладили на льду.
[35]Бактерии E.coli штамма Тор 10 были трансформированы 2 мкл лигазных смесей при помощи электропоратора MicroPulser (BioRad, #1652100), после чего были посеяны на селективную среду (LB-arap с 1% канамицином) в чашку Петри и инкубировались в течение ночи.
[36]Для выявления колоний, содержащих целевую плазмидную ДНК pCMV6_T7_synthUTR_AGN, был проведен ПЦР-скрининг колоний с использованием олигонуклеотидов pCMV6 w.i. for screening F последовательностью 5 '-TAAATGGCCCGCCTGGCATT-3' и pCMV6 w.i. for screening R MCS последовательностью 5'-GATCC AAGCTTGGGCCCGTAC-3'.
[37]Целевая плазмидная ДНК pCMV6_T7_synthUTR_AGN была выделена из E.coli, выращенной в 200 мл жидкой среды LB с добавлением 1% канамицина. Для выделения и очистки плазмидной ДНК использовали набор Plasmid-20-Maxi (Биолабмикс, Россия).
[38]Полученная в результате плазмида pCMV6_T7_synthUTR_AGN, имеющая нуклеотидную последовательность SEQ ID NO 1, содержит следующие элементы:
[39]- рекомбинантный промотор РНК-полимеразы фага Т7 с нуклеотидами AGN для встройки кэп-аналога и вспомогательной последовательности, размером 32 п. н.;
[40]- рекомбинантную 5'-нетранслируемую область 5'-UTR-4, необходимую для повышения стабильности мРНК и эффективностй трансляции целевого белка, размером 48 п. н.;
[41]- полилинкер MCS, содержащий сайты узнавания эндонуклеаз рестрикции EcoRI, Sail, Sfr274I, CciNI, PspLI, PspOMI, ApaI, HindIII, BamHI, размером 46 п. н., необходимый для встройки рамки считывания целевого гена;
[42]- рекомбинантную 3'-нетранслируемую область 3'-UTR AES-mtRNR1, необходимую для повышения стабильности мРНК и эффективности трансляции целевого белка, размером 278 п. н.;
[43]- сайт узнавания эндонуклеазы рестрикции XbaI, необходимый для линеаризации плазмидной ДНК перед ее использованием в реакции транскрипции in vitro в качестве матрицы, размером 6 п. н.;
[44]- последовательность CMV энхансера (380 п. н.), CMV промотора (204 п. н.), сигнала полиаденилирования hGH (623 п. н.), ориджины репликации ColE1 (589 п. н.) и фага fl (456 п. н.), сигнал полиаденилирования тимидинкиназы герпесвируса (48 п. н.), ген устойчивости к канамицину (795 п. н.) под промотором SV40 (358 п. н.), а также промотор AmpR (105 п. н.)
[45]Нуклеотидная последовательность плазмиды
[46]pCMV6_T7_synthUTR_AGN (SEQ ID NO 1) приведена в перечне последовательностей.
[47]Пример 2. Получение рекомбинантной плазмидной ДНК pCMV6_T7_synthUTR_GNN, содержащей в своем составе нетранслируемые области 5'-UTR-4 и 3'-UTR AES-mtRNR1.
[48]Получение рекомбинантной плазмидной ДНК
[49]pCMV6_T7_synthUTR_GNN (фиг.2) проводилось аналогично примеру 1, за исключением того, что в плазмиду pBluescript SK (+) по сайтам AbsI и BstXI (СибЭнзайм, Россия) была встроена последовательность промотора РНК полимеразы фага Т7 с нуклеотидами для встройки кэп-аналога 5'-GNN-3', где N - любой нуклеотид, и вспомогательной последовательностью 5'-АТААТ-3', 5'-UTR-4, MCS и 3'-UTR-AES-mtRNR1.
[50]Полученная в результате плазмида pCMV6_T7_synthUTR_GNN, имеющая нуклеотидную последовательность SEQ ID NO 2, содержит следующие элементы:
[51]- рекомбинантный промотор РНК-полимеразы фага Т7 с нуклеотидами GNN для встройки кэп-аналога и вспомогательной последовательности, размером 32 п. н.;
[52]рекомбинантную 5'-нетранс лиру ему ю область 5'-UTR-4, необходимую для повышения стабильности мРНК и эффективности трансляции целевого белка, размером 48 п. н.;
[53]- полилинкер MCS, содержащий сайты узнавания эндонуклеаз рестрикции EcoRI, Sail, Sfr274I, CciNI, PspLI, PspOMI, ApaI, HindIII, BamHI, размером 46 п. н., необходимый для встройки рамки считывания целевого гена;
[54]- рекомбинантную 3'-нетранслируемую область 3'-UTR AES-mtRNR1, необходимую для повышения стабильности мРНК и эффективности трансляции целевого белка, размером 278 п. н.;
[55]- сайт узнавания эндонуклеазы рестрикции XbaI, необходимый для линеаризации плазмидной ДНК перед ее использованием в реакции транскрипции in vitro в качестве матрицы, размером 6 п. н.;
[56]- последовательность CMV энхансера (380 п. н.), CMV промотора (204 п. н.), сигнала полиаденилирования hGH (623 п. н.), ориджины репликации ColEl (589 п. н.) и фага fl (456 п. н.), сигнал полиаденилирования тимидинкиназы герпесвируса (48 п. н.), ген устойчивости к канамицину (795 п. н.) под промотором SV40 (358 п. н.), а также промотор AmpR (105 п. н.).,
[57]Нуклеотидная последовательность плазмиды pCMV6_T7_synthUTR_GNN (SEQ ID NO 2) приведена в перечне последовательностей.
[58]Пример 3. Синтез полноразмерных мРНК Luc2 и hMGFP с 5'-UTR-4 и 3'-UTR AES-mtRNR1.
[59]Для получения полноразмерных РНК по матрице плазмид pCMV6_T7_synthUTR_AGN_Luc2, pCMV6_T7_synthUTR_AGN_hMGFP, pCMV6_T7_synthUTR_GNN_Luc2, pCMV6_T7_synthUTR_GNN_hMGFP, плазмиды линеаризовали с помощью эндонуклеазы рестрикции XbaI. Линеаризованные плазмиды использовали в реакциях Т7-транскрипции в качестве ДНК-матриц. Спустя 4 часа из реакционных смесей объемом 100 мкл отбирали аликвоты объемом 10 мкл для проверки целостности мРНК и их подвижности в 1.5%-ном агарозном геле. Электрофоретический анализ показал наличие в реакционных смесях целостных продуктов необходимой длины (фиг.3), выход реакций при этом составил -300 нг/мкл реакции.
[60]Таким образом, показано, что созданные плазмидные конструкции на основе векторов pCMV6_T7_synthUTR_AGN/GNN обеспечивают образование мРНК необходимой длины, что позволяет использовать их для высокоэффективной наработки мРНК.
[61]Пример 4. Трансфекция клеток НЕК293Т/17 мРНК Luc2 с дальнейшей детекцией люминесцентного сигнала.
[62]Реакционную смесь, содержащую мРНК Luc2 с комбинацией 5'-UTR-4 и 3'-UTR AES-mtRNR1 и кэп-аналогами ARC А или AG очищали с помощью сорбции на кремниевых колонках с дальнейшей высокоэффективной жидкостной хроматографией для удаления невключившихся трифосфатов и низкоорганических соединений. После очистки мРНК формировали липоплексы с молекулами-доставщиками 2x3:DOPE (1,26-бис(холест-5-ен-3|3-илоксикарбиламино)-7,11,16,20-
[63]тетраазогексакозан тетрагидрохлорид (2X3) в CHCl-СН3ОН (1:1 vol.) в смеси с раствором 1,2-диолеил-зп-глицеро-3-фосфоэтаноламина (DOPE) в CHCl3) в клеточной среде DMEM/F12 (Gibco, США) без добавления сыворотки. В качестве контроля формировали липоплексы с мРНК, содержащими 5'- и 3'-UTR β-глобина. Полученные липоплексы трансфицировали в клетки человека НЕК293Т/17 с дальнейшей детекцией специфического люминесцентного (фиг.4). Детекцию люминесцентного сигнала проводили с помощью планшетного ридера ClarioStar с оценкой относительного уровня люминесценции в режиме реального времени.
[64]Было показано, что 5'-UTR-4 и 3'-UTR AES-mtRNR1 в составе структуры мРНК Luc2 обеспечивают более высокий уровень специфического сигнала, а значит и более высокий уровень экспрессии люциферазы.
[65]Пример 5. Трансфекция клеток НЕК293Т/17 мРНК hMGFP с дальнейшей детекцией флюоресцентного сигнала.
[66]Реакционную смесь, содержащую мРНК hMGFP с комбинацией 5'-UTR-4 и 3'-UTR AES-mtRNR1 и кэп-аналогом AG очищали с помощью сорбции на кремниевых колонках с дальнейшей высокоэффективной жидкостной хроматографией для удаления невключившихся трифосфатов и низкоорганических соединений. После очистки мРНК формировали липоплексы с коммерческим трансфектантом Lipofectamine 3000 в клеточной среде DMEM/F12 (Gibco, США) без добавления сыворотки. В качестве контролей формировали липоплексы с мРНК, не содержащими нетранслируемых областей и с мРНК, содержащими 5'- и 3'-UTR β-глобина. Полученные липоплексы трансфицировали в клетки человека НЕК293Т/17 с дальнейшей детекцией специфического флюоресцентного сигнала (фиг.5, 6). Детекцию флюоресцентного сигнала проводили методом проточной цитометрии с оценкой относительного уровня флюоресценции и процента hMGFP-положительных клеток.
[67]Было показано, что 5'-UTR-4 и 3'-UTR AES-mtRNR1 в составе структуры мРНК hMGFP обеспечивают более высокий уровень специфического сигнала, а значит и более высокий уровень экспрессии зеленого флюоресцентного белка.
[68]Пример 6. Детекция специфического люминесцентного сигнала от мРНК Luc2 при введении in vivo модельным животным.
[69]10 мкг мРНК Luc2 с кэпом AG и с UTR β-глобина либо комбинацией 5'-UTR-4 и 3'-UTR AES-mtRNR1 вводили внутримышечно в составе липоплексов с молекулами-доставщиками 2x3:DOPE мышам линии Balb/c в возрасте 2х-4х недель. Спустя 8 ч, 24 ч, 32 ч, 48 ч, 72 ч и 96 ч проводили детекцию люминесцентного сигнала в режиме реального времени с помощью молекулярного имиджера IVIS Lumina Х5. Для этого мышам вводили 150 мкл D-люциферина в концентрации 24 мг/мл и спустя 15 минут проводили измерение сигнала. Мыши, инъецированные мРНК с разными комбинациями UTR демонстрировали продолжительный (более 4х дней) люминесцентный сигнал (фиг.7). При этом люминесцентный сигнал от мышей, инъецированных мРНК с 5'-UTR-4 и 3'-UTR AES-mtRNR1 (на фиг.7 - столбцы черного цвета) продемонстрировал увеличение интенсивности в 6 раз спустя 72 часа от введения препарата мРНК в отличие от мРНК с UTR β-глобина. Таким образом, включение в состав мРНК комбинации этих нетранслируемых областей значительно увеличивает экспрессию репортерной мРНК in vivo.
[71]1. Zhuang X, Qi Y, Wang M, et al. mRNA Vaccines Encoding the HA Protein of Influenza A H1N1 Virus Delivered by Cationic Lipid Nanoparticles Induce Protective Immune Responses in Mice. Vaccines (Basel). 2020; 8, 123, doi: 10.3390/vaccines8010123, PMID: 32164372.
[72]2. Panova, E.A., Kleymenov, D.A., Shcheblyakov, D.V., et al. Single-domain antibody delivery using an mRNA platform protects against lethal doses of botulinum neurotoxin A. Front. Immunol. 2023, 14, 1098302, 10.3389/fimmu.2023.1098302, PMID: 36865543.
[73]3. Linares-Femandez S, Moreno J, Lambert E, et al. Combining an optimized mRNA template with a double purification process allows strong expression of in vitro transcribed mRNA. Mol Ther Nucleic Acids. 2021;26:945-956. doi: 10.1016/j.omtn.2021.10.007, PMID: 34692232.
[74]4. OriGene URL: https://www.origene.com/catalog/vectors/mammalian-expression-vectors/ps100001/pcmv6-entry-mammalian-expression-vector (дата обращения: 07.05.2024).
[75]5. addgene URL: https://www.addgene.org/vector-database/7482/ (дата обращения: 14.05.24).
[76]6. Conrad, T., Plumbom, I., Alcobendas, M., Vidal R., Sauer S. Maximizing transcription of nucleic acids with efficient T7 promoters. Commun Biol. 2020, 3, 439, doi: 10.1038/s42003-020-01167-x, PMID: 32796901.
[77]7. Orlandini von Niessen, A.G., Poleganov, M.A., Rechner, C., Plaschke, A., et al. Improving mRNA-Based Therapeutic Gene Delivery by Expression- Augmenting 3' UTRs Identified by Cellular Library Screening. Molecular Therapy 2019, 27, 824-836, doi:10.1016/j.ymthe.2018.12.011, PMID: 30638957.
[79]<?xml version="1.0" encoding="UTF-8"?>
[80]<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
[81]1.3//EN" "ST26SequenceListing_V1_3.dtd">
[82]<ST26SequenceListing dtdVersion="V1_3"
[83]fileName="plasmidpatentUTR1.xml" softwareName="WIPO Sequence"
[84]softwareVersion="2.3.0" productionDate="2024-05-15">
[85] <ApplicationIdentification>
[86] <IPOfficeCode>RU</IPOfficeCode>
[87] <ApplicationNumberText>1</ApplicationNumberText>
[88] <FilingDate>2024-05-15</FilingDate>
[89] </ApplicationIdentification>
[90] <ApplicantFileReference>C:\Users\234\Desktop\plasmidpatentUTR</Applic
[92] <EarliestPriorityApplicationIdentification>
[93] <IPOfficeCode>RU</IPOfficeCode>
[94] <ApplicationNumberText>1</ApplicationNumberText>
[95] <FilingDate>2024-05-15</FilingDate>
[96] </EarliestPriorityApplicationIdentification>
[97] <ApplicantName languageCode="ru">Федеральное государственное
[98]бюджетное учреждение науки Институт химической биологии и
[99]фундаментальной медицины Сибирского отделения Российской академии
[100]наук (ИХБФМ СО РАН)</ApplicantName>
[101] <ApplicantNameLatin>The Institute of Chemical Biology and
[102]Fundamental Medicine of the Siberian Branch of the Russian Academy of
[103]Sciences </ApplicantNameLatin>
[104] <InventionTitle languageCode="ru">Рекомбинантные плазмидные ДНК
[105]pCMV6_T7_synthUTR_AGN/GNN, кодирующие искусственные матричные РНК,
[106]обеспечивающие синтез белков в клетках млекопитающих</InventionTitle>
[107] <SequenceTotalQuantity>2</SequenceTotalQuantity>
[108] <SequenceData sequenceIDNumber="1">
[110] <INSDSeq_length>5057</INSDSeq_length>
[111] <INSDSeq_moltype>DNA</INSDSeq_moltype>
[112] <INSDSeq_division>PAT</INSDSeq_division>
[113] <INSDSeq_feature-table>
[115] <INSDFeature_key>source</INSDFeature_key>
[116] <INSDFeature_location>1..5057</INSDFeature_location>
[117] <INSDFeature_quals>
[119] <INSDQualifier_name>mol_type</INSDQualifier_name>
[120] <INSDQualifier_value>other DNA</INSDQualifier_value>
[122] <INSDQualifier id="q2">
[123] <INSDQualifier_name>organism</INSDQualifier_name>
[124] <INSDQualifier_value>Homo sapiens</INSDQualifier_value>
[126] </INSDFeature_quals>
[128] </INSDSeq_feature-table>
[129] <INSDSeq_sequence>aacaaaatattaacgcttacaatttccattcgccattcaggctgcgcaa
[130]ctgttgggaagggcgatcggtgcgggcctcttcgctattacgccagctggcgaaagggggatgtgctgca
[131]aggcgattaagttgggtaacgccagggttttcccagtcacgacgttgtaaaacgacggccagtgccaagc
[132]tgatctatacattgaatcaatattggcaattagccatattagtcattggttatatagcataaatcaatat
[133]tggctattggccattgcatacgttgtatctatatcataatatgtacatttatattggctcatgtccaata
[134]tgaccgccatgttgacattgattattgactagttattaatagtaatcaattacggggtcattagttcata
[135]gcccatatatggagttccgcgttacataacttacggtaaatggcccgcctggctgaccgcccaacgaccc
[136]ccgcccattgacgtcaataatgacgtatgttcccatagtaacgccaatagggactttccattgacgtcaa
[137]tgggtggagtatttacggtaaactgcccacttggcagtacatcaagtgtatcatatgccaagtccgcccc
[138]ctattgacgtcaatgacggtaaatggcccgcctggcattatgcccagtacatgaccttacgggactttcc
[139]tacttggcagtacatctacgtattagtcatcgctattaccatggtgatgcggttttggcagtacaccaat
[140]gggcgtggatagcggtttgactcacggggatttccaagtctccaccccattgacgtcaatgggagtttgt
[141]tttggcaccaaaatcaacgggactttccaaaatgtcgtaataaccccgccccgttgacgcaaatgggcgg
[142]taggcgtgtacggtgggaggtctatataagcagagctcatgcagctaatacgactcactataaggataat
[143]ctgaaacacggtggagagtttattgcaaaataacgcgtccattcgacagaattcgtcgactcgagcggcc
[144]gcgtacgggcccaagcttggatccctggtactgcatgcacgcaatgctagctgcccctttcccgtcctgg
[145]gtaccccgagtctcccccgacctcgggtcccaggtatgctcccacctccacctgccccactcaccacctc
[146]tgctagttccagacacctcccaagcacgcagcaatgcagctcaaaacgcttagcctagccacacccccac
[147]gggaaacagcagtgattaacctttagcaataaacgaaagtttaactaagctatactaaccccagggttgg
[148]tcaatttcgtgccagccacacctctagacccgggtggcatccctgtgacccctccccagtgcctctcctg
[149]gccctggaagttgccactccagtgcccaccagccttgtcctaataaaattaagttgcatcattttgtctg
[150]actaggtgtccttctataatattatggggtggaggggggtggtatggagcaaggggcaagttgggaagac
[151]aacctgtagggcctgcggggtctattgggaaccaagctggagtgcagtggcacaatcttggctcactgca
[152]atctccgcctcctgggttcaagcgattctcctgcctcagcctcccgagttgttgggattccaggcatgca
[153]tgaccaggctcagctaatttttgtttttttggtagagacggggtttcaccatattggccaggctggtctc
[154]caactcctaatctcaggtgatctacccaccttggcctcccaaattgctgggattacaggcgtgaaccact
[155]gctcccttccctgtccttctgattttaaaataactataccagcaggaggacgtccagacacagcataggc
[156]tacctggccatgcccaaccggtgggacatttgagttgcttgcttggcactgtcctctcatgcgttgggtc
[157]cactcagtagatgcctgttgaattgggtacgcggccagcggcgagcggtatcagctcactcaaaggcggt
[158]aatacggttatccacagaatcaggggataacgcaggaaagaacatgtgagcaaaaggccagcaaaaggcc
[159]aggaaccgtaaaaaggccgcgttgctggcgtttttccataggctccgcccccctgacgagcatcacaaaa
[160]atcgacgctcaagtcagaggtggcgaaacccgacaggactataaagataccaggcgtttccccctggaag
[161]ctccctcgtgcgctctcctgttccgaccctgccgcttaccggatacctgtccgcctttctcccttcggga
[162]agcgtggcgctttctcatagctcacgctgtaggtatctcagttcggtgtaggtcgttcgctccaagctgg
[163]gctgtgtgcacgaaccccccgttcagcccgaccgctgcgccttatccggtaactatcgtcttgagtccaa
[164]cccggtaagacacgacttatcgccactggcagcagccactggtaacaggattagcagagcgaggtatgta
[165]ggcggtgctacagagttcttgaagtggtggcctaactacggctacactagaagaacagtatttggtatct
[166]gcgctctgctgaagccagttaccttcggaaaaagagttggtagctcttgatccggcaaacaaaccaccgc
[167]tggtagcggtggtttttttgtttgcaagcagcagattacgcgcagaaaaaaaggatctcaagaagatcct
[168]ttgatcttttctacggggtctgacgctcagtggaacgaaaactcacgttaagggattttggtcatgagat
[169]tatcaaaaaggatcttcacctagatccttttaaattaaaaatgaagttttaaatcaatctaaagtatata
[170]tgagtaacctgaggctatggcagggcctgccgccccgacgttggctgcgagccctgggccttcacccgaa
[171]cttggggggtggggtggggaaaaggaagaaacgcgggcgtattggccccaatggggtctcggtggggtat
[172]cgacagagtgccagccctgggaccgaaccccgcgtttatgaacaaacgacccaacaccgtgcgttttatt
[173]ctgtctttttattgccgtcatagcgcgggttccttccggtattgtctccttccgtgtttcagttagcctc
[174]cccctagggtgggcgaagaactccagcatgagatccccgcgctggaggatcatccagccggcgtcccgga
[175]aaacgattccgaagcccaacctttcatagaaggcggcggtggaatcgaaatctcgtgatggcaggttggg
[176]cgtcgcttggtcggtcatttcgaaccccagagtcccgctcagaagaactcgtcaagaaggcgatagaagg
[177]cgatgcgctgcgaatcgggagcggcgataccgtaaagcacgaggaagcggtcagcccattcgccgccaag
[178]ctcttcagcaatatcacgggtagccaacgctatgtcctgatagcgatccgccacacccagccggccacag
[179]tcgatgaatccagaaaagcggccattttccaccatgatattcggcaagcaggcatcgccatgggtcacga
[180]cgagatcctcgccgtcgggcatgctcgccttgagcctggcgaacagttcggctggcgcgagcccctgatg
[181]ctcttcgtccagatcatcctgatcgacaagaccggcttccatccgagtacgtgctcgctcgatgcgatgt
[182]ttcgcttggtggtcgaatgggcaggtagccggatcaagcgtatgcagccgccgcattgcatcagccatga
[183]tggatactttctcggcaggagcaaggtgagatgacaggagatcctgccccggcacttcgcccaatagcag
[184]ccagtcccttcccgcttcagtgacaacgtcgagcacagctgcgcaaggaacgcccgtcgtggccagccac
[185]gatagccgcgctgcctcgtcttgcagttcattcagggcaccggacaggtcggtcttgacaaaaagaaccg
[186]ggcgcccctgcgctgacagccggaacacggcggcatcagagcagccgattgtctgttgtgcccagtcata
[187]gccgaatagcctctccacccaagcggccggagaacctgcgtgcaatccatcttgttcaatcatgcgaaac
[188]gatcctcatcctgtctcttgatcgatctttgcaaaagcctaggcctccaaaaaagcctcctcactacttc
[189]tggaatagctcagaggccgaggcggcctcggcctctgcataaataaaaaaaattagtcagccatggggcg
[190]gagaatgggcggaactgggcggagttaggggcgggatgggcggagttaggggcgggactatggttgctga
[191]ctaattgagatgcatgctttgcatacttctgcctgctggggagcctggggactttccacacctggttgct
[192]gactaattgagatgcatgctttgcatacttctgcctgctggggagcctggggactttccacaccctaact
[193]gacacacattccacagctggttctttccgcctcaggactcttcctttttcaatattattgaagcatttat
[194]cagggttattgtctcatgagcggatacatatttgaatgtatttagaaaaataaacaaataggggttccgc
[195]gcacatttccccgaaaagtgccacctgacgcgccctgtagcggcgcattaagcgcggcgggtgtggtggt
[196]tacgcgcagcgtgaccgctacacttgccagcgccctagcgcccgctcctttcgctttcttcccttccttt
[197]ctcgccacgttcgccggctttccccgtcaagctctaaatcgggggctccctttagggttccgatttagtg
[198]ctttacggcacctcgaccccaaaaaacttgattagggtgatggttcacgtagtgggccatcgccctgata
[199]gacggtttttcgccctttgacgttggagtccacgttctttaatagtggactcttgttccaaactggaaca
[200]acactcaaccctatctcggtctattcttttgatttataagggattttgccgatttcggcctattggttaa
[201]aaaatgagctgatttaacaaaaatttaacgcgaatttt</INSDSeq_sequence>
[204] <SequenceData sequenceIDNumber="2">
[206] <INSDSeq_length>5057</INSDSeq_length>
[207] <INSDSeq_moltype>DNA</INSDSeq_moltype>
[208] <INSDSeq_division>PAT</INSDSeq_division>
[209] <INSDSeq_feature-table>
[211] <INSDFeature_key>source</INSDFeature_key>
[212] <INSDFeature_location>1..5057</INSDFeature_location>
[213] <INSDFeature_quals>
[215] <INSDQualifier_name>mol_type</INSDQualifier_name>
[216] <INSDQualifier_value>other DNA</INSDQualifier_value>
[218] <INSDQualifier id="q4">
[219] <INSDQualifier_name>organism</INSDQualifier_name>
[220] <INSDQualifier_value>Homo sapiens</INSDQualifier_value>
[222] </INSDFeature_quals>
[224] </INSDSeq_feature-table>
[225] <INSDSeq_sequence>aacaaaatattaacgcttacaatttccattcgccattcaggctgcgcaa
[226]ctgttgggaagggcgatcggtgcgggcctcttcgctattacgccagctggcgaaagggggatgtgctgca
[227]aggcgattaagttgggtaacgccagggttttcccagtcacgacgttgtaaaacgacggccagtgccaagc
[228]tgatctatacattgaatcaatattggcaattagccatattagtcattggttatatagcataaatcaatat
[229]tggctattggccattgcatacgttgtatctatatcataatatgtacatttatattggctcatgtccaata
[230]tgaccgccatgttgacattgattattgactagttattaatagtaatcaattacggggtcattagttcata
[231]gcccatatatggagttccgcgttacataacttacggtaaatggcccgcctggctgaccgcccaacgaccc
[232]ccgcccattgacgtcaataatgacgtatgttcccatagtaacgccaatagggactttccattgacgtcaa
[233]tgggtggagtatttacggtaaactgcccacttggcagtacatcaagtgtatcatatgccaagtccgcccc
[234]ctattgacgtcaatgacggtaaatggcccgcctggcattatgcccagtacatgaccttacgggactttcc
[235]tacttggcagtacatctacgtattagtcatcgctattaccatggtgatgcggttttggcagtacaccaat
[236]gggcgtggatagcggtttgactcacggggatttccaagtctccaccccattgacgtcaatgggagtttgt
[237]tttggcaccaaaatcaacgggactttccaaaatgtcgtaataaccccgccccgttgacgcaaatgggcgg
[238]taggcgtgtacggtgggaggtctatataagcagagctcatgcagctaatacgactcactataaggataat
[239]ctgaaacacggtggagagtttattgcaaaataacgcgtccattcgacagaattcgtcgactcgagcggcc
[240]gcgtacgggcccaagcttggatccctggtactgcatgcacgcaatgctagctgcccctttcccgtcctgg
[241]gtaccccgagtctcccccgacctcgggtcccaggtatgctcccacctccacctgccccactcaccacctc
[242]tgctagttccagacacctcccaagcacgcagcaatgcagctcaaaacgcttagcctagccacacccccac
[243]gggaaacagcagtgattaacctttagcaataaacgaaagtttaactaagctatactaaccccagggttgg
[244]tcaatttcgtgccagccacacctctagacccgggtggcatccctgtgacccctccccagtgcctctcctg
[245]gccctggaagttgccactccagtgcccaccagccttgtcctaataaaattaagttgcatcattttgtctg
[246]actaggtgtccttctataatattatggggtggaggggggtggtatggagcaaggggcaagttgggaagac
[247]aacctgtagggcctgcggggtctattgggaaccaagctggagtgcagtggcacaatcttggctcactgca
[248]atctccgcctcctgggttcaagcgattctcctgcctcagcctcccgagttgttgggattccaggcatgca
[249]tgaccaggctcagctaatttttgtttttttggtagagacggggtttcaccatattggccaggctggtctc
[250]caactcctaatctcaggtgatctacccaccttggcctcccaaattgctgggattacaggcgtgaaccact
[251]gctcccttccctgtccttctgattttaaaataactataccagcaggaggacgtccagacacagcataggc
[252]tacctggccatgcccaaccggtgggacatttgagttgcttgcttggcactgtcctctcatgcgttgggtc
[253]cactcagtagatgcctgttgaattgggtacgcggccagcggcgagcggtatcagctcactcaaaggcggt
[254]aatacggttatccacagaatcaggggataacgcaggaaagaacatgtgagcaaaaggccagcaaaaggcc
[255]aggaaccgtaaaaaggccgcgttgctggcgtttttccataggctccgcccccctgacgagcatcacaaaa
[256]atcgacgctcaagtcagaggtggcgaaacccgacaggactataaagataccaggcgtttccccctggaag
[257]ctccctcgtgcgctctcctgttccgaccctgccgcttaccggatacctgtccgcctttctcccttcggga
[258]agcgtggcgctttctcatagctcacgctgtaggtatctcagttcggtgtaggtcgttcgctccaagctgg
[259]gctgtgtgcacgaaccccccgttcagcccgaccgctgcgccttatccggtaactatcgtcttgagtccaa
[260]cccggtaagacacgacttatcgccactggcagcagccactggtaacaggattagcagagcgaggtatgta
[261]ggcggtgctacagagttcttgaagtggtggcctaactacggctacactagaagaacagtatttggtatct
[262]gcgctctgctgaagccagttaccttcggaaaaagagttggtagctcttgatccggcaaacaaaccaccgc
[263]tggtagcggtggtttttttgtttgcaagcagcagattacgcgcagaaaaaaaggatctcaagaagatcct
[264]ttgatcttttctacggggtctgacgctcagtggaacgaaaactcacgttaagggattttggtcatgagat
[265]tatcaaaaaggatcttcacctagatccttttaaattaaaaatgaagttttaaatcaatctaaagtatata
[266]tgagtaacctgaggctatggcagggcctgccgccccgacgttggctgcgagccctgggccttcacccgaa
[267]cttggggggtggggtggggaaaaggaagaaacgcgggcgtattggccccaatggggtctcggtggggtat
[268]cgacagagtgccagccctgggaccgaaccccgcgtttatgaacaaacgacccaacaccgtgcgttttatt
[269]ctgtctttttattgccgtcatagcgcgggttccttccggtattgtctccttccgtgtttcagttagcctc
[270]cccctagggtgggcgaagaactccagcatgagatccccgcgctggaggatcatccagccggcgtcccgga
[271]aaacgattccgaagcccaacctttcatagaaggcggcggtggaatcgaaatctcgtgatggcaggttggg
[272]cgtcgcttggtcggtcatttcgaaccccagagtcccgctcagaagaactcgtcaagaaggcgatagaagg
[273]cgatgcgctgcgaatcgggagcggcgataccgtaaagcacgaggaagcggtcagcccattcgccgccaag
[274]ctcttcagcaatatcacgggtagccaacgctatgtcctgatagcgatccgccacacccagccggccacag
[275]tcgatgaatccagaaaagcggccattttccaccatgatattcggcaagcaggcatcgccatgggtcacga
[276]cgagatcctcgccgtcgggcatgctcgccttgagcctggcgaacagttcggctggcgcgagcccctgatg
[277]ctcttcgtccagatcatcctgatcgacaagaccggcttccatccgagtacgtgctcgctcgatgcgatgt
[278]ttcgcttggtggtcgaatgggcaggtagccggatcaagcgtatgcagccgccgcattgcatcagccatga
[279]tggatactttctcggcaggagcaaggtgagatgacaggagatcctgccccggcacttcgcccaatagcag
[280]ccagtcccttcccgcttcagtgacaacgtcgagcacagctgcgcaaggaacgcccgtcgtggccagccac
[281]gatagccgcgctgcctcgtcttgcagttcattcagggcaccggacaggtcggtcttgacaaaaagaaccg
[282]ggcgcccctgcgctgacagccggaacacggcggcatcagagcagccgattgtctgttgtgcccagtcata
[283]gccgaatagcctctccacccaagcggccggagaacctgcgtgcaatccatcttgttcaatcatgcgaaac
[284]gatcctcatcctgtctcttgatcgatctttgcaaaagcctaggcctccaaaaaagcctcctcactacttc
[285]tggaatagctcagaggccgaggcggcctcggcctctgcataaataaaaaaaattagtcagccatggggcg
[286]gagaatgggcggaactgggcggagttaggggcgggatgggcggagttaggggcgggactatggttgctga
[287]ctaattgagatgcatgctttgcatacttctgcctgctggggagcctggggactttccacacctggttgct
[288]gactaattgagatgcatgctttgcatacttctgcctgctggggagcctggggactttccacaccctaact
[289]gacacacattccacagctggttctttccgcctcaggactcttcctttttcaatattattgaagcatttat
[290]cagggttattgtctcatgagcggatacatatttgaatgtatttagaaaaataaacaaataggggttccgc
[291]gcacatttccccgaaaagtgccacctgacgcgccctgtagcggcgcattaagcgcggcgggtgtggtggt
[292]tacgcgcagcgtgaccgctacacttgccagcgccctagcgcccgctcctttcgctttcttcccttccttt
[293]ctcgccacgttcgccggctttccccgtcaagctctaaatcgggggctccctttagggttccgatttagtg
[294]ctttacggcacctcgaccccaaaaaacttgattagggtgatggttcacgtagtgggccatcgccctgata
[295]gacggtttttcgccctttgacgttggagtccacgttctttaatagtggactcttgttccaaactggaaca
[296]acactcaaccctatctcggtctattcttttgatttataagggattttgccgatttcggcctattggttaa
[297]aaaatgagctgatttaacaaaaatttaacgcgaatttt</INSDSeq_sequence>
[300]</ST26SequenceListing>


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}