[2]Настоящее изобретение относится к микробиологической промышленности, а именно к получению продуцента изолимонной кислоты на основе дрожжей вида Yarrowia lipolytica, модифицированных методами генной инженерии.
[3]Предшествующий уровень техники
[4]Возрастающий интерес к продукции природного изомера изолимонной кислоты (трео-Ds(+)-изолимонная кислота) связан с использованием ее в фармацевтической и пищевой промышленностях [1].
[5]Перспективной является разработка технологии получения изолимонной кислоты с использованием микроорганизмов. Микробиологический синтез позволяет получить природный изомер и является сравнительно быстрым и простым способом, что делает продукт доступным и рентабельным для дальнейшего использования. Способность продуцировать изолимонную кислоту обнаружена у мицелиального гриба вида Penicillium purpurogenum, представителей дрожжей Candida ravautii, Candida catenulate, а также Yarrowia lipolytica [2]. Среди указанных микроорганизмов дрожжи Y. lipolytica являются наиболее физиологически и генетически изученными, для них разработаны технологии промышленной ферментации, системы генетической трансформации, подходы для экспрессии и инактивации генов, секвенирован геном [3].
[6]Известно, что изоцитрат (изолимонная кислота) синтезируется в митохондриях в процессе осуществления цикла Кребса посредством реакции изомеризации цитрата в изоцитрат, катализируемой ферментом аконитазой. В условиях избытка источника углерода и лимита источника азота происходит частичное торможение следующего этапа цикла Кребса - окислительного декарбоксилирования изоцитрата, катализируемого ферментом изоцитратдегидрогеназой (Idh), что приводит к накоплению интермедиантов цикла Кребса изоцитрата и цитрата в митохондриях [4]. Причина торможения декарбоксилирования изоцитрата заключается в том, что клетки начинают синтезировать ионы аммония из аденозинмонофосфата (АМФ) за счет повышения активности фермента аденозинмонофосфатдеаминазы (АМФД), при этом снижение АМФ, являющегося кофактором митохондриальной изоцитратдегидрогеназы, приводит к ингибированию этого фермента и, как следствие, к накоплению цитрата и изоцитрата [4].
[7]В условиях избытка источника углерода и лимита источника азота повышение экспрессии гена YlAMPD, ответственного за синтез аденозинмонофосфатдеаминазы в дрожжах Y. lipolytica, приводит к увеличению пула синтезируемого изоцитрата и цитрата. Так, природный штамм Y. lipolytica W29 за 5 дней культивирования в условиях избытка источника углерода и лимита по азоту продуцирует 38.5 г/л смеси цитрата и изоцитрата, а штамм Yarrowia lipolytica W29 YlAMPD, обладающий увеличенной экспрессией гена YlAMPD, в тех же условиях производит 47.5 г/л смеси цитрата и изоцитрата [5].
[8]Внутренняя мембрана митохондрии непроницаема для крупных молекул, таких как органические кислоты, аминокислоты, нуклеотиды, коферменты и кофакторы. Транспортную функцию таких молекул через мембрану осуществляет особая группа белков - митохондриальные транспортеры [6].
[9]Известно, что в дрожжах Y. lipolytica за транспорт цитрата в митохондрию и из нее отвечает белок YlYhm2p [5], а за транспортировку изоцитрата между митохондриями и цитоплазмой отвечает белок YlSfc1p [7].
[10]Инактивация гена YlYHM2 приводит к неспособности митохондрий клетки транспортировать цитрат из митохондрий в цитоплазму, тем самым позволяя полностью использовать цитрат внутри митохондрий для дальнейших биохимических превращений. Штамм Y. lipolytica W29 dYlyhm2, полученный из природного штамма Y. lipolytica W29 в результате делеции гена YlYHM2, в условиях избытка источника углерода и лимита по азоту накапливает в среде культивирования только 0.05 г/л цитрата за 5 дней культивирования, а исходный штамм - 30.1 г/л цитрата [5].
[11]В условиях избытка источника углерода и лимита источника азота повышение уровня экспрессии гена YlSFC1 приводит к увеличению продукции изолимонной кислоты и изменению соотношения изолимонная кислота / лимонная кислота. Штамм Υ. lipolytica W29 YlSFC1 за 144 ч культивирования в 1 л ферментере накапливает 69.4 г/л изолимонной кислоты, при этом соотношение концентраций изолимонной кислоты к лимонной кислоте составляет 2.3:1, для исходного штамма те же показатели составляют 7.5 г/л и 0.1:1, соответственно [7].
[12]Краткое описание изобретения
[13]Технической проблемой, на решение которой направлено настоящее изобретение является расширение арсенала микроорганизмов, способных продуцировать изолимонную кислоту.
[14]Техническим результатом заявляемого изобретения является рекомбинантный штамм дрожжей Yarrowia lipolytica 583 ВКПМ Y-4657 с инактивированным геном YlYHM2 и усиленной экспрессией генов YlAMPD и YlSFC1, продуцирующий изолимонную кислоту в условиях избытка источника углерода и лимита источника азота.
[15]Заявляемый штамм получен из родительского штамма Y. lipolytica W29 YlAMPD, обладающего усиленной экспрессией гена YlAMPD, путем следующих модификаций:
[16]- инактивация гена YlYHM2
[17]- усиление экспрессии гена YlSFC1
[18]Краткое описание рисунков
[19]Изобретение проиллюстрировано следующими фигурами графического изображения:
[20]Фиг. 1. Схема генетической конструкции pUC-f1YlYHM2-URA3
[21]Фиг. 2. Схема генетической конструкции pTEFin-uno-YlSFC1
[22]Пример 1. Конструирование заявляемого штамма
[23]Для проведения экспериментов используют штамм Yarrowia lipolytica W29 YlAMPD, который получен из штамма Yarrowia lipolytica W29 URA3- (ВКПМ Y-3372) путем трансформации экспрессионной кассетой, содержащей селективный маркер URA3, ограниченный с двух сторон сайтами lox66 и lox71, узнаваемыми рекомбиназой Cre, а также ген YlAMPD под контролем промотора hp4d и терминатора гена YlXPR2.
[24]1.1 Инактивация гена ΥlΥΗΜ2
[25]Известным методом [8] на базе штамма Y. lipolytica W29 YlAMPD получают ауксотрофный по урацилу штамм Y. lipolytica W29 YlAMPD URA3-.
[26]Генетическую конструкцию pUC-f1YlYHM2-URA3 [5], изображенную на фиг. 1 и содержащую следующие элементы, клонированные на векторе pUC19:
[27]- фланги YlSFC1up и YlSFC1down - участки, гомологичные к областям хромосомы штамма Yarrowia lipolytica W29 YlAMPD выше и ниже, относительно рамки гена YlSFC1,
[28]- селективный маркер URA3, ограниченный с двух сторон сайтами lox66 и lox71, узнаваемыми рекомбиназой Cre,
[29]обрабатывают эндонуклеазой рестрикции EheI, и 1 мкг полученной делеционной кассеты f1YlYHM2-URA3 используют для трансформации штамма Yarrowia lipolytica W29 YlAMPD URA3- методом электропорации [9]. Трансформанты селектируют на минимальной среде YNBD (Himedia Μ139 с добавлением глюкозы 20 г/л) по прототрофности к урацилу и далее, методом ПЦР отбирают клоны, которые содержат делецию гена ΥlΥΗΜ2, используя праймеры:
[30]CCCAGAACCTTCTTGCATTCCA (YlYhm2-conf-F)
[31]CCTCCCACATGAGTCATGCTTCC (YlYhm2-conf-R)
[32]В результате ПЦР с хромосомной ДНК клонов, содержащих делецию гена YlYHM2, получают продукт размером 3555 п. о. Один из отобранных клонов Y. lipolytica W29 YlAMPD dYlyhm2, содержащий делецию гена ΥlΥΗΜ2, используют в дальнейшей работе.
[33]1.2 Усиление экспрессии гена YlSFC1
[34]На базе штамма Y. lipolytica W29 YlAMPD dYlyhm2, аналогично примеру 1.1, получают ауксотрофный штамм Y. lipolytica YlAMPD dYlyhm2 URA3-.
[35]Генетическую конструкцию pTEFin-uno-YlSFC1 [7], изображенную на фиг .2 и содержащую следующие элементы, клонированные на векторе pUC19:
[36]- ген YlSFC1 под контролем промотора TEF1 и терминатора гена XPR2,
[37]- селективный маркер URA3, ограниченный с двух сторон сайтами lox66 и lox71, узнаваемыми рекомбиназой Cre,
[38]обрабатывают эндонуклеазами рестрикции SapI и EheI, и 2 мкг полученной экспрессионной кассеты TEFin-uno-YlSFC1 используют для трансформации штамма Y. lipolytica YlAMPD dYlyhm2 URA3- методом электропорации. Трансформанты отбирают на минимальной среде YNBD (Himedia M139 с добавлением глюкозы 20 г/л) по прототрофности к урацилу.
[39]Культивирование отобранных трансформантов проводят в трех независимых повторностях в пробирках (50 мл) с рабочим объемом 10 мл при 30°С и постоянном перемешивании (250 об/мин) в течение 5 суток в среде YNBD9N0.2 (среда Himedia M151 с добавлением глюкозы 90 г/л, (NH4)2SO4 2 г/л, 100 мМ калий-фосфатного буфера рН=6.8). В качестве контролей используют штамм Y. lipolytica W29 YlAMPD dYlyhm2 и природный штамм Y. lipolytica. На вторые сутки культивирования для поддержания рН добавляют мел 200 мг.Пробы отбирают на пятые сутки культивирования. К культуральной жидкости добавляют равный объем 1М HCl, центрифугируют 1 мин (14500 об/мин), супернатант переносят в новую микропробирку и делают необходимые разведения. Количество продуцируемых изолимонной и лимонной кислот в супернатанте определяют с помощью метода высокоэффективной жидкостной хроматографии.
[40]Лучший из отобранных трансформантов 583 имеет продукцию изолимонной кислоты 41.4 г/л. (табл.).
[41]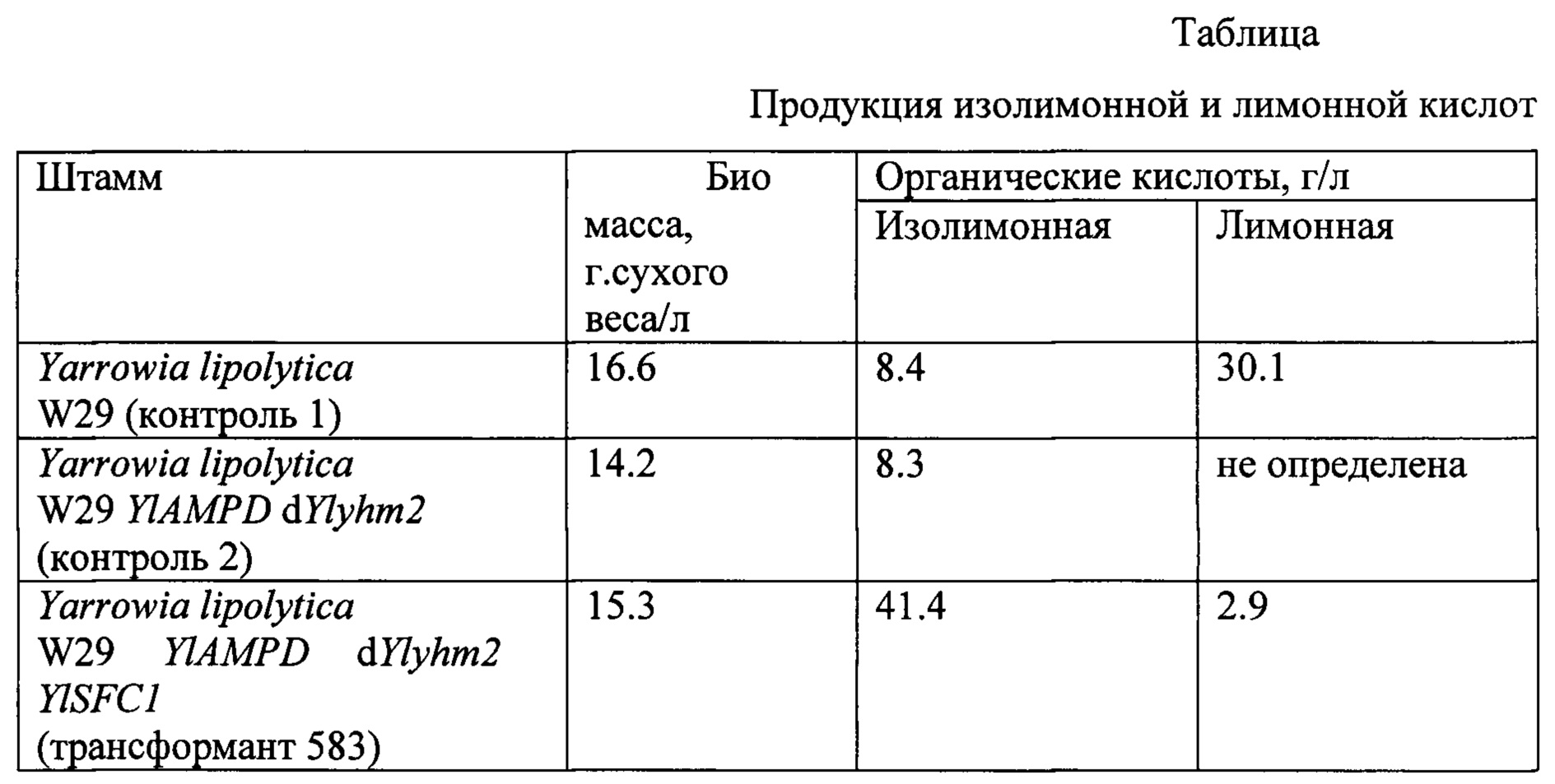
[42]Этот трансформант депонирован в БРЦ ВКПМ НИЦ «Курчатовский институт» - ГосНИИгенетика (117545 Москва, 1-ый Дорожный пр-д, д.1) как штамм Yarrowia lipolytica 583 ВКПМ Y-4657.
[43]Заявляемый штамм Yarrowia lipolytica 583 ВКПМ Y-4657 характеризуется следующими признаками:
[44]Культурально-морфологические признаки
[45]Суточная культура в жидкой среде YPD (мас. %: дрожжевой экстракт - 1, пептон - 2, глюкоза - 2, вода - остальное) представлена овальными, удлиненно-овальными, округлыми клетками размером 4.6-6.0×4.5-12.5 мкм. Почкование полярное или латеральное, на узком основании. К третьим суткам большинство клеток почкуются и образуют псевдомицелий. Колонии на агаризованной среде YPD кремового цвета, пастообразные, слегка приподнятые в центре, морщинистые, с фестончатым краем. Штрих на агаризованной среде YPD непрерывный, плоский, блестящий, кремово-белого цвета, пастообразный, края ровные, со временем становится складчатым. Спор не образует.
[46]Физиолого-биохимические признаки
[47]Облигатный аэроб. Сахара не сбраживает. Ассимилирует глюкозу, глицерин, этанол, ацетат, гидрофобные субстраты. Не ассимилирует: мальтозу, лактозу, целлобиозу, трегалозу, мелибиозу, раффинозу, мелицитозу, инулин, крахмал, D-ксилозу, L- и D-арабинозу, раммозу, дульцит, инозит, D-глюкозамин и глюкуроновую, 2-кетоглюконовую, 5-кетоглюконовую кислоты. Не ассимилирует нитраты. Не растет в безвитаминной среде, требует тиамин, не требует биотин. Оптимальное значение рН для роста 5,5-7,0. Не растет при 37°С. Максимальная температура роста 35°С.
[48]Пример 2. Получение изолимонной кислоты путем культивирования заявляемого штамма Y. lipolytica 583 ВКПМ Y-4657
[49]Культивирование штамма Y. lipolytica 583 ВКПМ Y-4657 проводят в 1-л ферментере Applikon BioBundle 1L с рабочим объемом 500 мл при температуре 29°С в минеральной среде следующего состава (мас. %): глюкоза - 4, (NH4)2SO4 - 0.3, MgSO4⋅7H2O - 0.14, NaCl - 0.05, Ca(NO3)2 - 0.08, KH2PO4 - 0.2, K2HPO4 - 0.02, биотин - 0.00005, тиамин - 0.001, вода - остальное. Концентрированный раствор микроэлементов следующего состава (мас. %): CuSO4⋅5H2O - 0,6, ΚΙ - 0.0088, MnSO4⋅5H2O - 0.3, Н3ВО3 - 0.02, CoCl2⋅6H2O - 0.0955, ZnSO4⋅7H2O - 4.2, FeSO4⋅7H2O - 6.5, H2SO4 - 0.5, вода - остальное готовят отдельно и вносят в количестве 4.6 мл на 1 л рабочего объема. В качестве пеногасителя используют адеканоль (0.1% об./об.). Каждые 24 ч в ферментер вносят подпитку глюкозы (30 г/л). Раствор 5М NaOH используют для поддержания значения рН среды равного 5 на протяжении первых суток культивирования, затем значение рН=6 до конца культивирования. Уровень растворенного в среде кислорода составляет не менее 50%.
[50]Подготовку посевной культуры проводят в два этапа:
[51]- пересев штамма на агаризованную среду YPD;
[52]- посев в жидкую среду YNBDbuf (среда Himedia Μ139 с добавлением глюкозы 20 г/л и 100 мМ калий-фосфатного буфера) в колбы с общим объемом 750 мл и рабочим объемом 70 мл и инкубация в течение 24 ч при 30°С и постоянном перемешивании (250 об/мин).
[53]Посевную культуру вносят в ферментер в количестве 10% от рабочего объема. Отбор проб осуществляют каждые 24 ч.
[54]Заявляемый штамм накапливает 136.7 г/л изолимонной кислоты за 166 ч культивирования, при этом соотношение изолимонная кислота / лимонная кислота достигает значения 7.4.
[55]Таким образом, заявляемый штамм Y. lipolytica 583 ВКПМ Y-4657 с инактивированным геном YlYHM2 и усиленной экспрессией генов YlAMPD и YlSFC1, в условиях избытка источника углерода и лимита источника азота продуцирует в значительном количестве целевой продукт изолимонную кислоту при пониженном содержании примеси лимонной кислоты.
[57]1. Aurich, Α., et al., Microbiologically produced carboxylic acids used as building blocks in organic synthesis. Subcell Biochem, 2012. 64: p. 391 - 423.
[58]2. Kamzolova, S.V., et al., Isocitric acid production from rapeseed oil by Yarrowia lipolytica yeast. Appl Microbiol Biotechnol, 2013. 97(20): p. 9133-9144.
[59]3. Nicaud, J.M., Yarrowia lipolytica. Yeast, 2012. 29(10): p. 409-418.
[60]4. Beopoulos, Α., J.M. Nicaud, and C. Gaillardin, An overview of lipid metabolism in yeasts and its impact on biotechnological processes. Appl Microbiol Biotechnol, 2011. 90(4): p. 1193-1206.
[61]5. Yuzbasheva, E.Y., et al., The mitochondrial citrate carrier in Yarrowia lipolytica: Its identification, characterization and functional significance for the production of citric acid. Metabolic Engineering, 2019. 54: p. 264-274.
[62]6. Palmieri, F., et al., Identification of mitochondrial carriers in Saccharomyces cerevisiae by transport assay of reconstituted recombinant proteins. Biochim Biophys Acta, 2006. 1757(9-10): p. 1249-1262.
[64]8. Yuzbasheva, E.Y., et al., Co-expression of glucose-6-phosphate dehydrogenase and acyl-CoA binding protein enhances lipid accumulation in the yeast Yarrowia lipolytica. N Biotechnol, 2017. 39: p. 18-21
[65]9. Wang, J.-H., W. Hung, and S.-H. Tsai, High efficiency transformation by electroporation of Yarrowia lipolytica. The Journal of Microbiology, 2011. 49(3): p. 469-472.

