TECHNICAL FIELD
[0001]The invention belongs to the field of biopharmaceutics, and particularly relates to read-through of nonsense mutation sites of monogenic hereditary diseases using a genetic code expanded non-natural amino acid system. Moreover, by modifying the tRNA of Methanosarcina barkeri (tRNAPyl), a new UAA and UGA-encoded non-natural amino acid system with high read-through efficiency was obtained, and the range of using the orthogonal pair of tRNAPyl and pyrrolysyl-tRNA synthetase (PylRS) was extended. It can be used to read through stop codons UAG, UAA and UGA of three nonsense mutations.
BACKGROUND
Hereditary Diseases Caused by Nonsense Mutations
[0002]There are many types of genetic mutations in the human genome, and nonsense mutations belong to one type of genetic mutations. Genetic mutations are heritable variations occured in genomic DNA molecules, including frameshift mutations and base substitutions. Frameshift mutations include insertions and deletions of bases, while base substitutions are mainly missense mutations and nonsense mutations. A nonsense mutation refers to the mutation of a certain base of the coding gene, resulting in stop codons UAG, UAA and UGA, and the stop codon does not encode any amino acid. The stop codon cannot be paired with an anticodon of a transfer RNA (tRNA), but can be recognized by a termination factor or a release factor, so as to terminate the synthesis of a peptide bond to terminate protein synthesis, and thus produces an incomplete and non-functional protein. The occurrence of nonsense mutations causes premature termination codons (PTC) in the gene box, which leads to two results of genetic coding, one produces a truncated protein and the other results in the decrease of the stability of the mRNA containing PTC, so as to leads to a nonsense-mediated mRNA degradation pathway (NMD). According to statistics, about 11.2% of hereditary diseases produce PTC mutations, called premature termination codons diseases (PTC diseases). On the other hand, many cancers also produce PTC mutations (KEELING K. M., WANG D., ONARD S. E., BEDWELL D. M. Suppression of premature termination codons as a therapeutic approach. Critical reviews in biochemistry and molecular biology, 2012, 47: 444-463.).
[0003]Duchenne muscular dystrophy (DMD) is a typical representative of PTC diseases. DMD is a serious muscle atrophy disease and the most common X-linked recessive hereditary disease. It is mainly characterized by progressivity and lethality. Nonsense mutations in the DMD gene are one of the main causes of DMD. Nonsense mutations produce premature termination codons UAG, UAA, UGA, resulting in a truncated polypeptide product that causes the patient to loss or lack functional dystrophin, which leads to muscle atrophy. According to reports, the incidence of Duchenne muscular dystrophy in live born baby boys is 1/6300 to 1/3500 [Dooley J, Gordon K E, Dodds L, MacSween J. Duchenne muscular dystrophy: a 30-year population-based Incidence study. Clin Pediatr (Phila), 2010, 49: 177-179.]. There is no effective method for curing this disease now. The onset of this disease mainly appears in childhood. It leads to loss of walking ability in adolescence, and early death in adulthood. It causes heavy psychological and economic burdens on patients, their families and the society.
[0004]Methods for the read-through of premature termination codons in previous studies include: (1) chemical small molecule-induced read-through: aminoglycosides such as G418 and non-aminoglycosides such as PTC124. In 1996, Howard et al. observed for the first time in the study of cystic fibrosis that aminoglycoside antibiotics can induce PTC read-through in mammalian cells to synthesize intact functional proteins. However, aminoglycoside antibiotics can cause serious adverse reactions while exerting nonsense inhibition, the most serious of which are ototoxicity and nephrotoxicity. And in February 2016, PTC124 was just rejected by the US FDA. (2) Exon skipping Method: Antisense nucleotide drugs for DMD patients who express proteins skipping exon 51. But the FDA has rejected Biomarin's drissapersen. Another company, Sarepta Therapeutics' Eteplirsen, will receive FDA's review results in May 2016; (3) Inhibitor tRNA read-through: Its anticodon loop is mutated and can be paired with a stop codon so that the stop codon can be read through. The main reason that this treatment is difficult to enter clinical applications is that the suppressor tRNA may recognize a normal stop codon, resulting in potential toxicity of an abnormal protein.
Genetic Code Expansion Technology
[0005]After several years of research, people have a comprehensive understanding of the translation mechanism of prokaryotic ribosomes. The crystal and electron microscopic structures of different ribosomes have been resolved, and the structures of most ammonia tRNA synthetases have also been obtained. Based on these findings, a technology of genetic code expansion, using amber stop codon (TAG) to encode a variety of non-natural amino acids and to make site-directed insertion in vivo, has been developed in recent years. To date, this technology has successfully make site-directed expression of several non-natural amino acids in the proteins of living cells, giving them novel physical, chemical and physiological properties. Using this method, non-natural amino acids (including affinity tags and photoisomerized amino acids, carbonyl amino acids, and glycosylated amino acids) can be introduced into proteins (L. Wang et al., (2001), SCIENCE 292: 498-500; J. W. Chin et al, 2002, Journal of the American Chemical Society 124: 9026-9027; J. W. Chin, & P. G. Schultz, 2002, Chem Bio Chem 11: 1135-1137). These studies indicate that it is possible and selective and routine to introduce chemical functional groups, for example, specific chemical groups such as carbonyl, alkynyl, and azido groups which generally effectively and selectively form stable covalent bonds, into proteins. After introduced into the pathogenic proteins, such groups can be used to study the mechanism of interaction between pathogenic proteins and other proteins.
[0006]After observing the crystal structure of the complex of tRNAPyl and PylRS synthetase, it is found that PylRS synthetase does not recognize the anticodon loop of tRNAPyl. Therefore, we believe that changing the base sequence of the anticodon loop of tRNAPyl does not affect the orthogonality of tRNAPyl and PylRS synthetase.
[0007]The non-natural aminoacyl tRNA synthetase is an aminoacyl tRNA synthetase from a microorganism such as archaea or Escherichia coli, which has been modified by positive and negative screening of artificial protein sequences and does not bind to endogenous tRNAs of E. coli or eukaryotes, and is an aminoacyl tRNA synthetase having specific sequence. Reference: Wang L, Schultz P G. Expanding the genetic code [J]. Angewandte chemie international edition, 2005, 44(1): 34-66.
[0008]The meaning of “orthogonality” of tRNA and non-natural aminoacyl tRNA synthetase as used herein means that this tRNA is not a substrate for any endogenous aminoacyl tRNA synthetase, and this aminoacyl tRNA synthetase cannot aminoacylate any endogenous tRNA. The members of this orthogonal pair have a unique correspondence with each other. The meaning of orthogonality can also be found in the reference: Wang L, Schultz P G. Expanding the genetic code [J]. Angewandte chemie international edition, 2005, 44(1): 34-66.
SUMMARY OF THE INVENTION
[0009]After considering and studying the prior art, the inventors have constructed PCMV-UUA (tRNAPylUUA/PylRS) and PCMV-UCA (tRNAPylUCA/PylRS) plasmids by modifying the tRNA of Methanosarcina barkeri (tRNAPyl) to obtain a new UAA and UGA-encoded non-natural amino acid system with high read-through efficiency, which can be used to read through the three stop codons UAG, UAA and UGA. The inventors have constructed a plasmid that mimics the endogenous premature termination codons—the introduction of premature termination codons on Smad gene consisting of introns and exons can be used to evaluate the efficiency of reading through the endogenous premature termination codons. The inventors have also used the genetic codon expansion technology to read through nonsense mutation sites in monogenic hereditary diseases and cancer suppressor genes in tumor cells to restore the expression of corresponding proteins.
[0010]The advantages of the invention may be embodied in one or more of the following:
[0011]1. A new UAA and UGA-encoded non-natural amino acid system with high read-through efficiency is obtained.
[0012]2. By using the genetic codon expansion technology, the read-through of nonsense mutations in hereditary diseases is realized, and the normal structures and functions of truncated proteins are restored.
[0013]In one aspect, the invention relates to a tRNA, wherein the base CUA on the anticodon loop of the tRNA is mutated to UUA or UCA, and the mutated tRNA can still be recognized by at least one non-natural aminoacyl tRNA synthetase which is orthogonal thereto.
[0014]In one aspect, the invention relates to a tRNA, wherein the anticodon loop of the tRNA is not bound to at least one non-natural aminoacyl tRNA synthetase which is orthogonal thereto.
[0015]In one aspect, the invention relates to a tRNA, wherein the tRNA is a tRNA derived from Methanosarcina barkeri (tRNAPyl), or a tRNA derived from E. coli, preferably, the tRNA is tRNAPylUUAhaving the sequence of SEQ ID NO: 1, or tRNAPylUCAhaving the sequence of SEQ ID NO: 2.
[0016]In one aspect, the invention relates to a non-natural amino acid system, wherein the system comprises the tRNA of any aspect of the invention and at least one non-natural aminoacyl tRNA synthetase which is orthogonal thereto or the encoding nucleic acid sequence thereof. Preferably, the non-natural amino acid system is selected from the group consisting of a lysine-like non-natural amino acid system, a leucine-like non-natural amino acid system, and a tyrosine-like non-natural amino acid system, optionally wherein the lysine-like non-natural amino acid system includes a tRNA derived from Methanosarcina barkeri (tRNAPyl) and pyrrolysyl-tRNA synthetase (PylRS), the leucine-like non-natural amino acid system includes a tRNA derived from E. coli (tRNALeu) and leucyl-tRNA synthetase (LeuRS), and the tyrosine-like non-natural amino acid system includes a tRNA derived from E. coli (tRNATyr) and tyrosyl-tRNA synthetase (TyrRS).
[0017]In one aspect, the invention relates to a non-natural amino acid system selected from the group consisting of:
[0018]Lys-diazirine (NAEK) as shown in
[0000]
[0019]Lys-azido as shown in
[0000]
[0020]at least one of other non-natural amino acids containing a diazirine or an azide structure, wherein the leucine-like non-natural amino acid is selected from the group consisting of Anap as shown in
[0000]
[0000]and the tyrosine-like non-natural amino acid is selected from pAcF as shown in
[0000]
[0021]In one aspect, the invention relates to a plasmid, a vector, a host cell or a kit comprising the tRNA of any aspect of the invention or a non-natural amino acid system of any aspect of the invention.
[0022]In one aspect, the invention relates to a method for genetic codon expansion, wherein the base CUA on the anticodon loop of a tRNA is point-mutated to UUA and UCA, and the mutated tRNA can still be recognized by its corresponding non-natural amino acid tRNA synthetase.
[0023]The method of any aspect of the invention, wherein the tRNA is a tRNA derived from Methanosarcina barkeri (tRNAPyl), or a tRNA derived from E. coli, preferably, the tRNA is tRNAPylUUAhaving the sequence of SEQ ID NO: 1, or tRNAPylUCAhaving the sequence of SEQ ID NO: 2.
[0024]In one aspect, the invention relates to use of the tRNA of any aspect of the invention or the non-natural amino acid system of any aspect of the invention, in the manufacture of a medicament for the treatment of a hereditary disease or cancer, wherein the hereditary disease or cancer is caused by a nonsense mutation in a gene. Preferably, the hereditary disease or cancer is caused by a nonsense mutation occurred in Dystrophin protein, tumor suppressor gene STK11 or EPHB2 protein.
[0025]The use of any aspect of the invention, wherein the hereditary disease and cancer are selected from the group consisting of: Duchenne muscular dystrophy, cystic fibrosis, hemophilia A, hemophilia B, lipid storage, ataxia telangiectasia, Hurler's syndrome, amaurotic familial idiocy, stomach cancer, and lung cancer.
[0026]In one aspect, the invention relates to a method for restoring normal expression and function of a nonsense mutant protein by read-through, wherein the tRNA of any aspect of the invention or the non-natural amino acid system of any aspect of the invention is introduced into a cell or an organism comprising a nonsense mutant protein.
[0027]A method of any aspect of the invention, wherein the introduced tRNA or non-natural amino acid system recognizes a nonsense mutation of the protein of interest and introduces a non-natural amino acid at a corresponding site of the nonsense mutation to allow the translation of the protein of interest to avoid premature termination and to synthesize an intact functional protein.
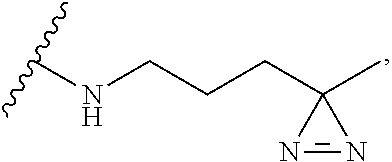
[0028]A method according to any aspect of the invention, wherein the introduced non-natural amino acid is Lys-diazirine at position N, and the manner for linking it in the protein is as follows:
[0000]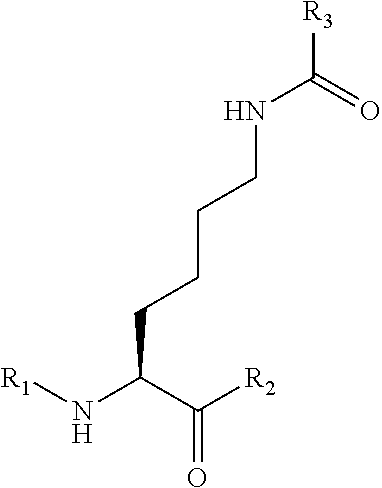
[0029]wherein the direction from R1 to R2 is the direction of the amino acid sequence from N-terminus to C-terminus, and position N may be an amino acid at any position of the pathogenic protein or the tumor suppressor gene protein, and correspondingly, R1 is an amino acid residue from position 1 to position N-1, R2 is an amino acid residue from position N+1 to the C-terminus, R3 is
[0000]
[0030]the introduced non-natural amino acid is Lys-azido at position N, and the manner for linking it in the pathogenic protein or the tumor suppressor gene protein is as follows:
[0000]
[0031]wherein the direction from R1 to R2 is the direction of the amino acid sequence from N-terminus to C-terminus, and position N may be any position of the pathogenic protein or the tumor suppressor gene protein according to claim 1, and correspondingly, R1 is an amino acid residue from position 1 to position N-1, R2 is an amino acid residue from position N+1 to the C-terminus,
R4 is
[0032]
[0033]In one aspect, the invention relates to a mammalian stable cell line HEK293-PYL, deposited on Nov. 17, 2015 under accession number CGMCC No. 11592.
[0034]A method for evaluating a genetic codon expansion technology, characterized in that the read-through efficiency thereof is evaluated by the amount of Smad protein expressed with the endogenous premature termination codon plasmid, preferably by the following steps:
[0035](1) cloning Smad gene into pcDNA3, the sequence of Smad gene preferably being set forth in SEQ ID NO:3;
[0036](2) mutating the codons at positions 39, 122, and 133 to UAG amber stop codon to obtain the mutant plasmids pcDNA3-Smad-39TAG, pcDNA3-Smad-122TAG and pcDNA3-Smad-133TAG, the mutant sequence preferably being set forth in SEQ ID NOs: 4-6;
[0037](3) transfecting the stable cell line HEK293-PYL with the mutant plasmids, adding the non-natural amino acid, culturing for 1-96 hours, preferably 12-72 hours, and most preferably 48 hours, then extracting the protein, detecting the full-length Smad protein by western blot, and evaluating the read-through efficiency according to the amount of the expressed full-length Smad protein.
[0038]In one aspect, the invention relates to a pair of primers, wherein said pair of primers have the following sequences:
[0000] | PCMV-UAG-UAA-for: |
| TGTAGATCGAATGGACTTTAAATCCGTTCAGCCGG |
| and |
| |
| PCMV-UAG-UAA-rev: |
| CCGGCTGAACGGATTTAAAGTCCATTCGATCTACA |
| or |
| |
| PCMV-UAG-UGA-for: |
| CATGTAGATCGAATGGACTTCAAATCCGTTCAGCCGGGTT |
| and |
| |
| PCMV-UAG-UGA-rev: |
| AACCCGGCTGAACGGATTTGAAGTCCATTCGATCTACATG. |
[0039]In one aspect, the invention relates to a method for restoring normal expression and function of a pathogenic protein in a monogenic hereditary disease and a tumor suppression gene protein in a tumor cell by read-through, which utilizes an optimized genetic codon expansion technology to insert a non-natural amino acid at a premature termination codon of a nonsense mutant protein.
[0040]In one aspect, the invention relates to tRNAs of Methanosarcina barkeri (tRNAPyl), wherein tRNAPylUUAand tRNAPylUCAare resulted from site-directed mutations of the anticodon loop of the original tRNAPylCUA, and the sequences thereof correspond to SEQ ID NO: 1 and SEQ ID NO: 2 respectively, and characterized in that they are perfectly paired with the stop codons UAA and UGA respectively. tRNAPylUUAand tRNAPylUCAare constructed on the PCMV-UUA and PCMV-UCA plasmids respectively.
[0041]In one aspect, the invention relates to a pathogenic protein or a tumor suppressor gene protein, wherein the inserted non-natural amino acid is Lys-diazirine at position N, and the manner for linking it in the protein is as follows:
[0000]
[0042]wherein the direction from R1 to R2 is the direction of the amino acid sequence from N-terminus to C-terminus, and position N may be an amino acid at any position of the pathogenic protein or the tumor suppressor gene protein, and correspondingly, R1 is an amino acid residue from position 1 to position N-1, R2 is an amino acid residue from position N+1 to the C-terminus, R3 is
[0000]
[0043]In one aspect, the invention relates to a pathogenic protein or a tumor suppressor gene protein, wherein the introduced non-natural amino acid is Lys-azido at position N, and the manner for linking it in the pathogenic protein or the tumor suppressor gene protein is as follows:
[0000]
[0044]wherein the direction from R1 to R2 is the direction of the amino acid sequence from N-terminus to C-terminus, and position N may be any position of the pathogenic protein or the tumor suppressor gene protein according to claim 1, and correspondingly, R1 is an amino acid residue from position 1 to position N-1, R2 is an amino acid residue from position N+1 to the C-terminus,
[0000]
[0046]In one aspect, the invention relates to a genetic codon expansion technology, wherein the read-through efficiency thereof is evaluated by the amount of Smad protein expressed with the endogenous premature termination codon plasmid pcDNA3-Smad by the following steps:
[0047](1) cloning Smad gene having the original sequence of SEQ ID NO:3 into pcDNA3;
[0048](2) mutating the codons at positions 39, 122, and 133 to UAG amber stop codon to obtain the mutant plasmids pcDNA3-Smad-39TAG, pcDNA3-Smad-122TAG and pcDNA3-Smad-133TAG having the sequences set forth in SEQ ID NOs: 4-6;
[0049](3) transfecting the stable cell line HEK293-PYL with the mutant plasmids, adding the non-natural amino acids, culturing for 48 hours, then extracting the protein, detecting the full-length Smad protein by western blot.
[0050]In one aspect, the invention relates to a mammalian stable cell line stably expressing tRNA (tRNAPylCUA) and pyrrolysyl-tRNA synthetase (PylRS), which is HEK293-PYL, deposited on Nov. 17, 2015 under accession number CGMCC No. 11592, as well as a HEK293-PYL-TAA stable cell line stably expressing tRNAPylUUA/PylRS, and a HEK293-PYL-TGA stable cell line stably expressing tRNAPylUCA/PylRS.
DETAILED DESCRIPTION
[0051]Specifically, in a specific embodiment of the invention, three tRNAPyl/PylRS plasmids recognizing three stop codons (amber, ocher, opal) were constructed, and restored the expression of the DMD disease protein Dystrophin and read through the endogenous premature termination codon in the stable cell line HEK293-PYL (which was deposited in China General Microbiological Culture Collection Center (CGMCC) on Nov. 17, 2015 under accession number CGMCC No. 11592 with the classification name human HEK293T cell), and restored the expression of the tumor suppressor genes STK11 and EPHB2 proteins in the A549 and DU145 tumor cell lines. The following six steps were mainly involved: (1) PCMV-UUA (tRNAPylUUA/PylRS) and PCMV-UCA (tRNAPylUCA/PylRS) plasmids were constructed; (2) GFP reporter genes pcDNA3.1-GFP-39TAG; pcDNA3.1-GFP-39TAA; and pcDNA3.1-GFP-39TGA comprising premature termination codons were constructed; (3) According to nonsense mutation sites of DMD patients, Dp71b protein plasmids Dp71b3116TAG, Dp71b3317TAG, and Dp71b3601TAG comprising the premature termination codon UAG were constructed by introducing the premature termination codon into the corresponding sites of the isoform protein of dystrophin protein, Dp71b by the point mutation technology; (4) the plasmids pcDNA3-Smad-39TAG, pcDNA3-Smad-122TAG and pcDNA3-Smad-133TAG mimicking the endogenous premature termination codon were constructed by introducing the premature termination codon TAG into Smad gene (consisting of introns and exons); (5) The plasmids of step (1) and (2) were correspondingly cross-transfected into 293T cells; non-natural amino acids were added, and the green fluorescence was observed after culturing for 48 hours to compare the read-through efficiencies of the three stop codons; (6) The plasmids of step (3) were transfected into the stable cell line HEK293-PYL; non-natural amino acids were added and the protein was extracted after culturing for 48 hours; the Dp71b full-length protein was detected by western blot to show the restoration of the expression of the disease protein; 7) The plasmids of step (4) were transfected into the stable cell line HEK293-PYL; non-natural amino acids were added and the protein was extracted after culturing for 48 hours, and Smad full-length protein was detected by western blot to prove that the codon expansion technology can effectively inhibit the nonsense-mediated mRNA degradation pathway and read through the endogenous premature termination codons at different positions; (8) The tumor cell lines A549 and DU145 were transfected with PCMV-CUA(tRNAPylCUA/PylRS); non-natural amino acids were added and the protein was extracted after culturing for 48 hours to prove the restoration of expression of STK11 protein and the full-length EPHB2 protein in tumor cell lines A549 and DU145 by western blot.
[0052]In a specific embodiment of the present invention, point mutation primers were designed using PCMV-CUA (tRNAPylCUA/PylRS) as a template plasmid. The base CUA on the anticodon loop of tRNAPylCUA was mutated to UUA and UCA with the above primers using a site-directed mutagenesis kit to obtain PCMV-UUA (tRNAPylUUA/PylRS) and PCMV-UCA (tRNAPylUCA/PylRS) plasmids.
[0053]In a specific embodiment of the invention, the read-through efficiencies of three tRNAPylCUA/UUA/UCA/PylRSs were detected with GFP green fluorescent protein containing a premature termination codon. In the first step, the amino acid codon at position 39 of GFP fluorescent gene was point mutated to the three premature termination codons, UAG, UAA and UGA respectively by point mutation technology to obtain the three plasmids, pcDNA3.1-GFP-39TAG, pcDNA3.1-GFP-39TAA and pcDNA3.1-GFP-39TGA. In the second step, 293T cells were correspondingly crossly co-transfected with PCMV-CUA/UUA/CUA and pcDNA3.1-GFP-39TAG/TAAA/TGA. In the third step, green fluorescence was observed by fluorescence microscopy after adding non-natural amino acids and culturing for 48 hours. It was finally confirmed that tRNAPyl/PylRS had an efficient read-through effect on the stop codons perfectly paired thereto, among which the read-through efficiency for UAG was the highest, that for UGA was the second, and that for UAA was the lowest.
[0054]In a specific embodiment of the invention, the genetic codon expansion technology is applied to restore the expression of a nonsense mutant protein associated with a human hereditary disease. According to the position of the nonsense mutation in a human DMD disease, a point mutation was performed at the corresponding position of the wild-type Dp71b sequence to construct Dp71b protein plasmids Dp71b3116TAG (c.9346C>T, Dp71b3317TAG (c.9952C>T), and Dp71b3601TAG (c.10801C>T) containing the premature termination codon UAG. The plasmids were transfected into the stable cell line HEK293-PYL, and the protein was extracted after adding non-natural amino acids and culturing for 48 hours. The full-length Dp71b protein was detected by western blot, and the expression of disease protein was restored.
[0055]In a specific embodiment of the invention, the stable cell line HEK293-PYL is used to verify that tRNAPylCUA/PylRS read through the endogenous premature termination codons at different positions. In the first step, Smad gene consisting of introns and exons was cloned into the pcDNA3 plasmid. Then the amino acid codons at positions 39, 122 and 133 of Smad were mutated to the UAG premature termination codon by point mutation process to obtain plasmids pcDNA3-Smad-39TAG, pcDNA3-Smad-122TAG and pcDNA3-Smad-133TAG. The stable cell line comprising the tRNA of Methanosarcina barkeri (tRNAPyl) and pyrrolysyl-tRNA synthetase (PylRS) were transfected with the endogenous premature termination codon plasmid (pcDNA3-Smad-39TAG, pcDNA3-Smad-122TAG or pcDNA3-Smad-133TAG). The protein was extracted after adding non-natural amino acids and culturing for 48 hours. The full-length Smad protein was detected by western blot in all the three groups. It is proved that the genetic codon expansion technology can effectively inhibit the nonsense-mediated mRNA degradation pathway, read through the endogenous premature termination codons at different positions, and restore the expression of the full-length protein.
[0056]In a specific embodiment of the invention, the genetic codon expansion technology was used to read through a nonsense mutation site of a tumor suppressor gene in a tumor cell. PCMV-CUA(tRNAPylCUA/PylRS) was transfected into tumor cell lines A549 and DU145 (the nonsense mutation c.109C>T, p.Q37X occurred in STK11 on human lung cancer cell A 549 genome is the stop codon UAG; the nonsense mutation c.2167C>T., p.Q723X occurred in EPHB2 gene on the human prostate cancer cell DU 145 genome is the stop codon UAG). The protein was extracted after adding non-natural amino acids and culturing for 48 hours. The restoration of the expression of the full-length STK11 protein and the full-length EPHB2 protein in tumor cell lines A549 and DU145 by the genetic codon expansion technology was proved by western blot.
[0057]More specifically, the present invention provides
[0058]1. A method for restoring normal expression and function of a pathogenic protein in a monogenic hereditary disease and a tumor suppression gene protein in a tumor cell by read-through, which utilizes the genetic codon expansion technology to insert a non-natural amino acid at a premature termination codon of a nonsense mutant protein.
[0059]2. The genetic codon expansion technology according to item 1, consisting of a tRNA derived from Methanosarcina barkeri (tRNAPyl), pyrrolysyl-tRNA synthetase (PylRS) and non-natural amino acids, wherein said non-natural amino acid is selected from:
[0060]Lys-diazirine (NAEK) as shown in
[0000]
[0061]Lys-azido as shown in
[0000]
[0062]at least one of other non-natural amino acids containing a diazirine or an azide structure.
[0063]3. The tRNAs of Methanosarcina barkeri (tRNAPyl) according to item 2, wherein tRNAPylUUAand tRNAPylUCAare engineered from site-directed mutations of the anticodon loop of the original tRNAPylCUA, and the sequences thereof correspond to SEQ ID NO: 1 and SEQ ID NO: 2 respectively, and characterized in that they are perfectly paired with the stop codons UAA and UGA respectively. tRNAPylUUAand tRNAPylUCAare constructed on the PCMV-UUA and PCMV-UCA plasmids respectively.
[0064]4. A pathogenic protein or a tumor suppressor gene protein according to item 1, wherein the inserted non-natural amino acid is Lys-diazirine at position N, and the manner for linking it in the protein is as follows:
[0000]
[0065]5. wherein the direction from R1 to R2 is the direction of the amino acid sequence from N-terminus to C-terminus, and position N may be an amino acid at any position of the pathogenic protein or the tumor suppressor gene protein, and correspondingly, R1 is an amino acid residue from position 1 to position N-1, R2 is an amino acid residue from position N+1 to the C-terminus, R3 is
[0000]
[0066]6. A pathogenic protein or a tumor suppressor gene protein according to item 1, wherein the introduced non-natural amino acid is Lys-azido at position N, and the manner for linking it in the pathogenic protein or the tumor suppressor gene protein is as follows:
[0000]
[0067]wherein the direction from R1 to R2 is the direction of the amino acid sequence from N-terminus to C-terminus, and position N may be any position of the pathogenic protein or the tumor suppressor gene protein according to claim 1, and correspondingly, R1 is an amino acid residue from position 1 to position N-1, R2 is an amino acid residue from position N+1 to the C-terminus,
[0000]
[0069]7. The genetic codon expansion technology according to items 1-5, wherein the read-through efficiency thereof is evaluated by the amount of Smad protein expressed with the endogenous premature termination codon plasmid pcDNA3-Smad by the following steps:
[0070](1) cloning Smad gene having the original sequence of SEQ ID NO:3 into pcDNA3;
[0071](2) mutating the codons at positions 39, 122, and 133 to UAG amber stop codon to obtain the mutant plasmids pcDNA3-Smad-39TAG, pcDNA3-Smad-122TAG and pcDNA3-Smad-133TAG having the sequences set forth in SEQ ID NOs: 4-6;
[0072](3) transfecting the stable cell line HEK293-PYL with the mutant plasmids, adding the non-natural amino acids, culturing for 48 hours, then extracting the protein, detecting the full-length Smad protein by western blot.
[0073]8. A mammalian stable cell line stably expressing tRNA (tRNAPylCUA) and pyrrolysyl-tRNA synthetase (PylRS), which is HEK293-PYL, deposited on Nov. 17, 2015 under accession number CGMCC No. 11592.
BRIEF DESCRIPTION OF THE DRAWINGS
[0074]FIG. 1. tRNAPylCUA/PylRS, tRNAPylUUA/PylRS and tRNAPylUCA/PylRS read through GFP green fluorescent protein TAG, TAA and TGA stop codons respectively.
[0075]FIG. 2A. Establishment of the screening method for the stable cell line HEK293-PYL of orthogonal tRNA/aminoacyl tRNA synthetase.
[0076]FIG. 2B. Construction of the double virus overexpression system.
[0077]FIG. 2C. Construction of the pXH-12t-zeo vector.
[0078]FIG. 3. Western blot confirms that in the stable cell line HEK293-PYL, the expression of the protein is restored by reading through the premature termination codons on the disease protein dystrophin.
[0079]FIG. 4. Read-through of the endogenous premature termination codons in the stable cell line HEK293-PYL.
[0080]FIG. 5. Genetic codon expansion technology reads through the premature termination codons in A549 and DU145 tumor cell lines to restore the expression of STK11 and EPHB2 proteins.
[0081]What have been described above are only some embodiments of the invention. It will be apparent to those skilled in the art that various variations and modifications can be made without departing from the spirit and scope of the invention, which all fall into the protection scope of the present invention.
[0082]In order to better understand the present invention, the inventors have described and illustrated the specific experiments by the Examples, which are intended to illustrate and not to limit the scope of the present invention. Any variations or embodiments equivalent to the invention are included in the invention.
Example 1
Construction of PCMV-UUA (tRNAPylUUA/PylRS) and PCMV-UCA tRNAPylUCA/PylRS Plasmids
[0083](1) Preparation of Methanosarcina barkeri PCMV-CUA Plasmid (tRNAPylCUA/PylRS)
[0084]Plasmid pACYC-tRNA/PylRS (hereinafter referred to as PCMV-CUA) was obtained from Escherichia coli pACYC-tRNA/PylRS comprising pACYC-tRNA/PylRS, which was deposited on Jun. 14, 2011 under accession number CGMCC No: 4951 with the classification name Escherichia coli, which was obtained from the depository, China General Microbiological Culture Collection Center, Institute of Microbiology Chinese Academy of Sciences NO. 1 West Beichen Road, Chaoyang District, Beijing. The plasmid can express the tRNA synthetase (PylRS) which specifically recognizes the non-natural amino acids Lys-diazirine and Lys-azido and the tRNA which specifically recognizes the amber stop codon UAG (tRNAPylCUA).
[0085](2) Construction of PCMV-UUA (tRNAPylUUA/PylRS) and PCMV-UCA (tRNAPylUCA/PylRS) Plasmids by Point Mutation tRNAPylCUA
[0086]The inventors designed mutant primers for the anticodon loop of the mutant tRNAPylCUA. The specific primers are shown below.
[0000] |
| PCMV-UAG-UAA- | TGTAGATCGAATGGACTTTAAATCCGTTCAGCCGG |
| for | |
|
| PCMV-UAG-UAA- | CCGGCTGAACGGATTTAAAGTCCATTCGATCTACA |
| rev | |
|
| PCMV-UAG-UGA- | CATGTAGATCGAATGGACTTCAAATCCGTTCAGCC |
| for | GGGTT |
|
| PCMV-UAG-UGA- | AACCCGGCTGAACGGATTTGAAGTCCATTCGATCT |
| rev | ACATG |
|
[0087]Plasmids PCMV-UUA (tRNAPylUUA/PylRS) and PCMV-UCA (tRNAPylUCA/PylRS) were obtained by using PCMV-CUA as the template plasmid, point mutating the base CUA on the anticodon loop of tRNAPylCUA to UUA and UCA with the above primers using the site-directed mutagenesis kit (QuikChange® Lightning Site-Directed Mutagenesis Kits, Catalog #210518) according to the instructions. The mutation was verified to be successful by sequencing. The sequence of tRNAPylUUAis represented by SEQ ID NO: 1; the sequence of tRNAPylUCAis represented by SEQ ID NO: 2.
Example 2: Detection of Read-Through Efficiency of Three tRNAPylCUA/UUA/UCA/PylRS Orthogonal Systems Using GFP Green Fluorescent Protein Comprising Premature Termination Codons
[0088](1) Synthesis and Identification of the Non-Natural Amino Acid Lys-Diazirine
[0089]The chemical synthesis reaction scheme of the non-natural amino acid Lys-diazirine was as follows:
[0000]
[0090]As shown in the above scheme, 15 mL of the starting material 1 (5-hydroxy-2-pentanone) and 40 mL of liquid ammonia were stirred and reacted at −40° C. for 5 h. Then the temperature was lowered to −60° C. The solution of NH2OSO3H in methanol was slowly added dropwise. The mixture was allowed to warm to room temperature and allowed to react overnight. The precipitate was filtered off, and triethylamine was added to the supernatant. I2 was slowly added under ice bath until the color of the reaction solution became dark, and no bubbles were generated. After the reaction was completed, the solvent was evaporated, and the mixture was extracted with diethyl ether and dried. Ether was distilled off, and the remaining liquid was evaporated under reduced pressure to give 25.4 g of colorless viscous liquid product 2.
[0091]The above product 2 was dissolved in pyridine. 11 g of TsCl was added with stirring at 0° C. to react overnight. After the reaction was completed, the reaction mixture was poured into a mixture of concentrated hydrochloric acid and ice water, and extracted with ethyl ether. The ether layer was washed with 1N hydrochloric acid and 1N NaOH. The organic phase was dried to give 11.8 g of a colorless viscous liquid product 3.
[0092]The above product 3 was dissolved in DMF, and NaN3 was added to react at room temperature overnight until the reaction was completed. A lot of water was added, and the mixture was extracted with ethyl ether. Ethyl ether was distilled off, and the remaining product was mixed with THF:water (9:1). Triphenylphosphine was added and reacted at room temperature. After the completion of the reaction, 1N HCl was added and the mixture was stirred, and THF was spin dried. The unreacted starting materials, PPh3 and O═PPh3 were washed away with methylene chloride, and the mixture was adjusted to pH 12 with 1N NaOH. 4.0 g of product 4 was obtained after extracted with dichloromethane.
[0093]5.2 g of the starting material 5 (Boc-Lys-OMe) was reacted with carbonyldiimidazole to prepare 5.9 g of compound 6. Compound 6 was then coupled with the above product 4 (4.0 g) to give compound 7, which was finally deprotected in two steps to remove Boc and methyl ester to give desired 4.5 g product 8, Lys-diazirine. The result verified by spectrometry was:
[0094]1H NMR (400 MHz, D2O): δ 3.10 (1H, t, J=6.3 Hz), 2.96 (4H, m), 1.25 (10H, m), 0.90 (3H, s); 13C NMR (100 MHz, D2O): 183.63, 160.66, 56.00, 39.80, 39.30, 34.49, 30.84, 29.20, 26.75, 23.92, 22.43, 18.80; HREIMS m/z 308.16937 [M+1]+ (calcd for C12H22N5NaO3, 308.16931). It proved that the obtained Lys-diazirine structure was correct.
[0095](2) Construction of a GFP Reporter Gene Containing Premature Termination Codons
[0096]Green fluorescent protein GFP is the most commonly used reporter gene and a powerful tool for indicating the insertion of non-natural amino acids. It consists of 238 amino acids and its gene sequence is represented by SEQ ID NO: 7.
[0097]The GFP sequence was inserted into the pcDNA3.1 commercial plasmid, and the amino acid codon at position 39 of the GFP fluorescent gene was mutated to three premature termination codons UAG, UAA and UGA respectively. Primers capable of mutating the codon encoding the amino acid into three stop codons respectively were designed, and the specific primers are shown in the following table.
[0000] |
| GFP-39- | GGCGAGGGCGATGCCACCTAGGGCAAGCTGACCCTGAAGTTC |
| UAG-for | |
|
| GFP-39- | GAACTTCAGGGTCAGCTTGCCCTAGGTGGCATCGCCCTCGCC |
| UAG-for | |
|
| GFP-39- | GGCGAGGGCGATGCCACCTAAGGCAAGCTGACCCTGAAGTTC |
| UAA-for | |
|
| GFP-39- | GAACTTCAGGGTCAGCTTGCCTTAGGTGGCATCGCCCTCGCC |
| UAA-for | |
|
| GFP-39- | GGCGAGGGCGATGCCACCTGAGGCAAGCTGACCCTGAAGTTC |
| UAG-for | |
|
| GFP-39- | GAACTTCAGGGTCAGCTTGCCTCAGGTGGCATCGCCCTCGCC |
| UAG-for |
|
[0098]The expression plasmids (pcDNA3.1-GFP-39TAG, pcDNA3.1-GFP-39TAA and pcDNA3.1-GFP-39TGA) were constructed by using the wild-type GFP expression vector pcDNA3.1-GFP-WT as a template, mutating the amino acid codon at position 39 to three stop codons respectively with the site-directed mutagenesis kit (QuikChange® Lightning Site-Directed Mutagenesis Kits, Catalog #210518) according to the instructions. The mutation was verified to be successful by sequencing.
[0099](3) Verification of the Read-Through Efficiency of the Orthogonal System after Mutation by Transient Transfection of PCMV and pcDNA3.1-GFP Plasmids in 293T Cells
[0100]The pcDNA3.1-GFP obtained in step 2 of Example 2, and the PCMV plasmid of step 2 of Example 1 were mixed in a ratio of 1:2 according to the grouping of Table 3, and then mixed with the transfection reagent megatrans1.0 in a ratio of 1:3. They were added together to 293T cells. After 6 hours, the solution was changed, and NAEK was added at the concentration of 1 mM. The cells were further cultured in an incubator at 37° C., 5% CO2 for 48 hours. Then green fluorescence was observed by fluorescent microscopy. The result was shown in FIG. 1. It was finally confirmed that tRNAPyl/PylRS has an efficient read-through effect on the stop codon perfectly paired thereto, in which the read-through efficiency for UAG is the highest, that for UGA is the second, and that for UAA is the lowest.
[0000] |
| PCMV PLASMID AND GFP PLASMID GROUPING MIX |
| group | plasmid |
| |
| 1 | PCMV-TAG and pcDNA3.1-GFP-39TAG |
| 2 | PCMV-TAA and pcDNA3.1-GFP-39TAG |
| 3 | PCMV-TGA and pcDNA3.1-GFP-39TAG |
| 4 | PCMV-TAG and pcDNA3.1-GFP-39TAA |
| 5 | PCMV-TAA and pcDNA3.1-GFP-39TAA |
| 6 | PCMV-TGA and pcDNA3.1-GFP-39TAA |
| 7 | PCMV-TAG and pcDNA3.1-GFP-39TGA |
| 8 | PCMV-TAA and pcDNA3.1-GFP-39TGA |
| 9 | PCMV-TGA and pcDNA3.1-GFP-39TGA |
| |
Example 3: Reading Trough the Disease Protein Dystrophin in Three HEK293-Pyl Stable Cell Lines
[0101](1) Construction of the Stable Cell Line HEK293-PYL
[0102]Two lentiviral overexpression vectors carrying puromycin and hygromycin resistances were constructed, which respectively carry an aminoacyl tRNA synthetase and a reporter gene GFP with TAG mutation at position 39. The stable cell strain pylRS/GFP39TAG was obtained after two rounds of transfection of HEK-293T cells with viruses and screening with puromycin/hygromycin. Subsequently, three pXH-zeo-12tRNA vectors carrying 12 copies of tRNA (CUA\UUA\UCA) and zeomycin resistance were constructed. The cell strain pylRS/GFP39TAG was transfected with linearized plasmids, and then screened in the presence of UAA. Finally, GFP-positive cells were isolated (the cells were green in the presence of UAA, and were colorless in the absence of UAA) to obtain three HEK293-PYL stable cell lines expressing tRNAPylCUA/PylRS, tRNAPylUUA/PylRS and tRNAPylUCA/PylRS respectively (FIG. 2A).
A. Construction of the Vector
[0103]We firstly constructed two lentiviral overexpression vectors respectively carrying puromycin and hygromycin resistances, which respectively carry an aminoacyl tRNA synthetase and a reporter gene GFP with TAG mutation at position 39. See FIG. 2B. Starting from the psd31 vector, we firstly replaced sv40-puroR gene with IRES-puroR and IRES-hygroR genes respectively by BamHI/xbaI restriction enzyme cutting site, thus obtained two viral vectors psd31-IRES-puroR and psd31-IRES-hygroR with different resistances, wherein IRES was an internal ribosome entry site and was often used for polycistronic gene expression. For example, an IRES sequence was inserted after the gene of interest, followed by a selectable marker gene, such that the transcribed mRNA could simultaneously express two proteins. There were two advantages to overexpress the gene of interest using the IRES system: 1. The gene of interest shared a promoter with the marker gene, avoiding the occurrence of false positive; 2. The translation efficiency of IRES was lower than that of the traditional translation initiation site, so that the expression level of the gene of interest was higher than that of the marker gene. Therefore, we introduced the CMV-pylRS sequence and the CMV-GFP39TAG sequence respectively before the IRES site by the BamHI restriction enzyme cutting site, so that obtained the double-virus system psd31-CMV-pylRS-IRES-puroR/psd31-CMV-GFP39TAG-IRES-hygroR which could simultaneously overexpress two proteins of interest. The main primers used are shown in Table 4.
[0000] |
| SOE | IRES-hygro-for(BamHI) | CGGGATCCAATTCCGCCCCTCTC |
| PCR | IRES-hygro-middle-for: | CCCACAAGGAGACGACCTTCCATGAAAAAGCC |
| primers | | TGAACTCACC |
| IRES-hygro-middle- | GGTGAGTTCAGGCTTTTTCATGGAAGGTCGTC |
| rev: | TCCTTGTGGG |
| IRES-hygro-rev(xbaI): | GCTCTAGA TCATTCCTTTGCCCTCGGAC |
|
| SOE | 3.1-CMV-for(BamHI) | CGGGATCCGTTGACATTGATTATTGAC |
| PCR | CMV-GFP-middle-for: | CCCAAGCTGGCTAGTTAAGCTTGCCACCATGG |
| primers | | ATTACAAGGATGACGACG |
| CMV-GFP-middle-rev: | CGTCGTCATCCTTGTAATCCATGGTGGCAAGC |
| | TTAACTAGCCAGCTTGGG |
| GFP-his-rev(BamHI): | CGGGATCCTCAATGGTGATGGTGATGATG |
|
| PCR | Pro-P1-for(BamHI): | TGGATCCCCAATATTGGCCATTAGCC |
| primers | MbpyIRS-rev(bamHI): | TGGATCCAAAAATTATAGATTGGTTG |
|
| Sequencing | PSD31-BamHI- | CAGGGACAGCAGAGATCCAG |
| primers | sequencing-for: | |
| 31-IRES-BamHI-rev: | GGCTTCGGCCAGTAACGTTAG |
|
[0104]The inventors overexpressed the tRNA by means of plasmid stable transfection. In order to ensure the expression level of the tRNA, the inventors constructed the vector pXH-12t-zeo, the sequence of which is shown in SEQ ID NO: 8. (FIG. 2C)
B. Packaging and Transduction of the Lentivirus
[0105]The psd31-CMV-pylRS-IRES-puroR virus was first packaged and transfected into HEK293T cells. The screening concentration of puromycin was 0.6 ug/ml. After the stable cell line No. 1 was obtained, the psd31-CMV-pylRS-IRES-puroR virus was added. The screening concentration of hygromycin was 200 ug/ml. The stable cell line No. 2 was obtained.
C. Stable Transfection of the Plasmid
[0106]The inventors performed a third round of screening by stable plasmid transfection, and finally obtained a special cell line stably expressing orthogonal tRNA/aminoacyl tRNA synthetase. The steps were as follows:
[0107]A. After pXH-12t-zeo vector was linearized by restriction enzyme cutting, the stable cell line No. 2 expressing pylRS and GFP39TAG proteins was transfected (10 cm culture dish, 10 ug plasmid per dish, no antibiotics when being transfected).
[0108]B. After 6 hours of transfection, the solution was changed and non-natural amino acids were added.
[0109]C. After 48 hours of transfection, green fluorescence was observed, and the solution was changed, and 400 ug/ml of zeomycin was added.
[0110]D. The solution was changed every 3 days until all the cells of the blank group died, and the transfection group formed clones.
[0111]E. The GFP-positive clones were isolated and purified, and the culture was further expanded with half-dosage of zeomycin to obtain a 12t-zeo stable cell line HEK293-PYL.
[0112]The main points of screening for monoclones by plasmid stable transfection are as follows:
[0113]A. The cell density of the cells stably transfected by the plasmid is important. The cell density is sparse at the time of screening, and it is easy to die and difficult form a clone.
[0114]B. From the time of monoclonalization, it is necessary to increase the nutrients, serum and growth factors.
[0115]C. When the number of cells inoculated into the well as a monoclone is small, the signal between the cells becomes weak and the positive cells may be in poor condition or even die. A special culture solution can be used: at the cell confluence of 80%, the old culture solution is sterilized by a filter, and is mixed with the fresh culture solution at a ratio of 1:1 for use. Alternatively, increase the concentration of the serum suitably.
[0116]D. After the digestion of the monoclone, do not add zeomycin and UAA, and should add them after cell adhesion to avoid cell death.
[0117](2) Construction of the Dp71b Mutant Plasmid Containing the Premature Termination Codon UAG
[0118]The Dp71b sequence of the isoform of the Dystrophin protein is shown in SEQ ID NO: 9. The inventors performed point mutations on the wild-type Dp71b sequence according to the sites of nonsense mutations in Duchenne muscular dystrophy patients, and introduced the premature termination codon at different positions to construct Dp71b plasmids Dp71b3116TAG (c.9346C>T), Dp71b3317TAG (c.9952C>T) and Dp71b3601TAG(c.10801C>T) comprising the premature termination codon UAG, which are shown in SEQ ID NOs: 10 to 12. The mutation was verified to be successful by sequencing.
[0000] |
| Dp71b- | TGAAACTCCGAAGACTGTAGAAGGCCCTTTGCTTG |
| 9346-for | |
| |
| Dp71b- | CAAGCAAAGGGCCTTCTACAGTCTTCGGAGTTTCA |
| 9346-for | |
| |
| Dp71b- | CATCAGGCCAAATGTAACATCTGCAAATAGTGTCC |
| 9952-for | AATCATT |
| |
| Dp71b- | AATGATTGGACACTATTTGCAGATGTTACATTTGG |
| 9952-for | CCTGATG |
| |
| Dp71b- | GCTGGAGCAACCCTAGGCAGAGGCCAA |
| 10801-for | |
| |
| Dp71b- | TTGGCCTCTGCCTAGGGTTGCTCCAGC |
| 10801-for |
| |
[0119](2) Reading Through the Disease Protein Dystrophin in the Stable Cell Line HEK293-PYL
[0120]The Dp71b3116TAG, Dp71b3317TAG and Dp71b3601TAG plasmids obtained in step 2 of Example 3 were mixed with the transfection reagent megatrans1.0 in a ratio of 1:3, and were added together to the stable cell line HEK293-PYL. After 6 hours, the solution was changed and NAEK at 1 mM was added. After the cells were cultured in an incubator at 37° C., 5% CO2 for 48 hours, the protein was extracted. The production of the full-length dystrophin protein was detected by Western blot (the primary antibody was anti-dystrophin, which was a C-terminal antibody of an anti-dystrophin protein, catalog No. 12715-1-AP), as shown in FIG. 3. It was proved that tRNAPyl/PylRS could read through the premature termination codons at different positions and restore the expression of disease proteins.
Example 4: Investigation of the Effect of Reading Through the Endogenous Premature Termination Codon in the Stable Cell Line HEK293-Pyl
[0121](1) Construction of the Endogenous Premature Termination Codon Plasmids pcDNA3.1-Smad-39TAG; pcDNA3.1-Smad-39TAA; pcDNA3.1-Smad-39TGA
[0122]Smad gene sequence consisting of introns and exons (as shown in SEQ ID: 3) was inserted into the pcDNA3.1 commercial plasmid, and then the amino acid codons at positions 39, 122 and 133 of Smad were mutated to the premature termination codon UAG to obtain plasmids pcDNA3-Smad-39TAG, pcDNA3-Smad-122TAG and pcDNA3-Smad-133TAG (as shown in SEQ ID NOs: 4 to 6).
[0123](2) Verification of the Read-Through of the Endogenous Premature Termination Codon in the Stable Cell Line
[0124]The pcDNA3-Smad-39TAG, pcDNA3-Smad-122TAG or pcDNA3-Smad-133TAG plasmid obtained in step 1 of Example 4 was mixed with the transfection reagent megatrans1.0 in a ratio of 1:3, and was added to the stable cell line HEK293-PYL. After 6 hours, the solution was changed and NAEK at 1 mM was added. After the cells were cultured in an incubator at 37° C., 5% CO2 for 48 hours, the protein was extracted. The production of the full-length Smad protein was detected by Western blot (the primary antibody was anti-myc, which was a tag antibody), as shown in FIG. 4. It was verified that the genetic codon expansion technology could inhibit the nonsense-mediated mRNA degradation process, and read through the premature termination codon to restore the expression of the protein.
Example 5: Genetic Codon Expansion Reads Through Premature Termination Codon in the Genome of a Tumor Cell Line
[0125]According to the literature, STK11 on human lung cancer cell A 549 genome has a nonsense mutation, c.109C>T, p. Q37X, which is an amber stop codon UAG; EPHB2 gene on human prostate cancer cell DU 145 genome has a nonsense mutation, c.2167C>T, p. Q723X, which is an amber stop codon UAG.
[0126]The PCMV-CUA (tRNAPylCUA/PylRS) plasmid was mixed with the transfection reagent megatrans1.0 in a ratio of 1:3, and was transfected into A 549 and DU145 cells respectively. After 6 hours, the solution was changed and NAEK at 1 mM was added. After the cells were cultured in an incubator at 37° C., 5% CO2 for 48 hours, the protein was extracted. The production of the full-length STK11 and EPHB2 proteins was detected by Western blot (the primary antibodies were anti-STK11 and anti-EPHB2 respectively), as shown in FIG. 5. It was verified that the genetic codon expansion technology could read through the premature termination codon on the endogenous genome to restore the expression of the tumor suppressor gene proteins.