[1]Изобретение относится к биотехнологии и может быть использовано в сельском хозяйстве для повышения продуктивности растений картофеля и/или их защиты от действия абиотических стрессоров, в частности хлоридного засоления при выращивании на открытом грунте или закрытом грунте.
[2]Одной из причин сокращения площади земель, доступных для сельского хозяйства, является засоление. Выращивание сельскохозяйственных культур на засоленных территориях приводит к снижению их продуктивности, что, в свою очередь, сопровождается серьезными экономическими потерями. Применение соединений гормональной природы для повышения продуктивности растений в условиях абиотического стресса является эффективной и безопасной технологией. Стероидные гормоны демонстрируют протекторные свойства в условиях хлоридного засоления. Брассиностероиды с лактоновым кольцом, к которым относится 24-эпибрассинолид (ЭБЛ), являются наиболее активными и применяются для создания агропрепаратов. Ранее показана эффективность фунгицидной композиции, главным составляющим которой является ЭБЛ, при обработке вегетирующих растений и семян (Пат. BY 022248).
[3]Известен способ повышения роста и развития растений комплексным препаратом для предпосевной обработки семян на основе брассиностероидов. В состав препарата входят: брассинолид 250-350 мг/л, гумат 8-12 г/л, кальциевый бентонит 35-45 г/л, автолизат пивных дрожжей 10-14 г/л. Изобретение позволяет ускорить прорастание семян яровой пшеницы и развитие проростков, в частности, приводит к достоверному увеличению их длины (Пат. RU 2728677).
[4]Основной недостаток способа заключается в необходимости использования ряда химических соединений, в том числе брассинолида, в относительно высокой концентрации, что сопровождается увеличением финансовых затрат при выращивании растений.
[5]Известен способ повышения устойчивости растений эпибрассинолидом. Технология предполагает предпосевную обработку семян томатов в течение 10-12 часов раствором фитопрепарата, в его состав включен эпибрассинолид в концентрации 10 нМ/л, с последующей вторичной обработкой молодых растений раствором эпибрассинолида в концентрации 10 мкМ/л при норме расхода 2-3 мл/растение. Изобретение позволяет стимулировать рост и развитие растений, усиливать процессы генеративного развития и повышать продуктивность на фоне стрессовых условий (Пат. RU 2349088). Показано увеличение количества плодов томатов и ускорение наступления фазы бутонизации.
[6]Основной недостаток способа - вторичная обработка растений фитопрепаратом, гормон в котором находится в высокой концентрации, что приводит к повышенному расходу эпибрассинолида, и также способствует удорожанию конечного продукта.
[7]Ранее был разработан способ повышения устойчивости растений рапса к интенсивному хлоридному засолению, включающий культивирование растений рапса в условиях хлоридного засоления и обработку растений раствором биологически активного вещества, отличающийся тем, что растения рапса культивируют на питательной среде Хогланда-Снейдера в течение трех недель, после чего растения подвергают засолению хлоридом натрия при концентрации приблизительно 175 мМ, во время которого в раствор двукратно вносят 24-эпибрассинолид, в первый день засоления и через 7 суток после засоления (Пат. RU 2522519).
[8]Основными недостатками способа являются двукратное внесение активного вещества, кроме того, концентрация используемого стрессора слишком высока, что не соответствует реальному засолению пахотных земель. Засоленными признаются почвы, в которых электропроводность составляет и 4 дСм/м или более, что эквивалентно раствору 40 мМ NaCl.
[9]Биологически активные вещества (БАВ) - это соединения, которые способны оказывать выраженное воздействие на процессы жизнедеятельности даже при использовании в низких концентрациях. В предложенном способе в качестве такового выступает 24-эпибрассинолид, который является биологически активным веществом, фитостимулятором, регулирующим рост растений, ингибирующим транспирацию и снижающим увядание, а также средством, усиливающим холодостойкость, ускоряющим рост, утолщение или созревание плодов, корней и корнеплодов.
[10]Технической задачей изобретения является разработка экономичного способа повышения продуктивности растений картофеля, как в обычных условиях выращивания, так и при действии хлоридного засоления.
[11]Технический результат заключается в увеличении количества ярусов и биомассы растений картофеля, содержания в них фото синтетических пигментов, снижении уровня окислительного стресса при последующем действии солевого стресса.
[12]Задача решается однократным добавлением экологически безопасного фитогормона 24-эпибрассинолида в концентрации 10 нМ или 1000 нМ в питательную среду Мурасиге-Скуга на этапе трехнедельного клонирования растений картофеля in vitro.
[13]Для решения поставленной задачи предложен способ повышения продуктивности растений картофеля эпибрассинолидом в оптимальных условиях и при хлоридном засолении, включающий микроклональное размножение in vitro и выращивание растений картофеля на питательной среде Мурасиге-Скуга с обработкой растений картофеля раствором биологически активного вещества, в качестве которого добавляют 24-эпибрассинолид, отличающийся тем, что на начальном этапе трехнедельного (21 день) клонирования растений картофеля in vitro и до воздействия солевого стресса однократно добавляют 24-эпибрассинолид в концентрации 10 нМ или 1000 нМ, биологически активное вещество добавляют в питательную среду.
[14]Далее растения картофеля адаптировали к жидкой питательной среде. Ростовые показатели (длина побега и корня) фиксировали сразу после переноса растений из условий in vitro, затем через 7 и 14 суток. После адаптации к жидкой среде растения подвергались стрессу, вызванному избыточной концентрацией хлорида натрия (75 мМ NaCl) на протяжении 5 суток. Брассиностероды в условиях стресса вызывали отложенное ростостимулирующее действие на рост побега. Кроме того, защитный эффект ЭБЛ проявлялся в сохранении биомассы растений и количества ярусов. Известно, что продуктивность растения определяется не только его массой, но и фотосинтетической активностью его листьев, которая зависит от содержания фото синтетических пигментов. Отмечено поддержание уровня пигментов (хлорофилла  и каротиноидов) на уровне контрольных значений для растений, выращенных с добавлением ЭБЛ в питательную среду. Так же для обработанных гормоном растений отмечено снижение величины перекисного окисления липидов и эндогенного уровня пролина по сравнению с необработанными растениями.
и каротиноидов) на уровне контрольных значений для растений, выращенных с добавлением ЭБЛ в питательную среду. Так же для обработанных гормоном растений отмечено снижение величины перекисного окисления липидов и эндогенного уровня пролина по сравнению с необработанными растениями.
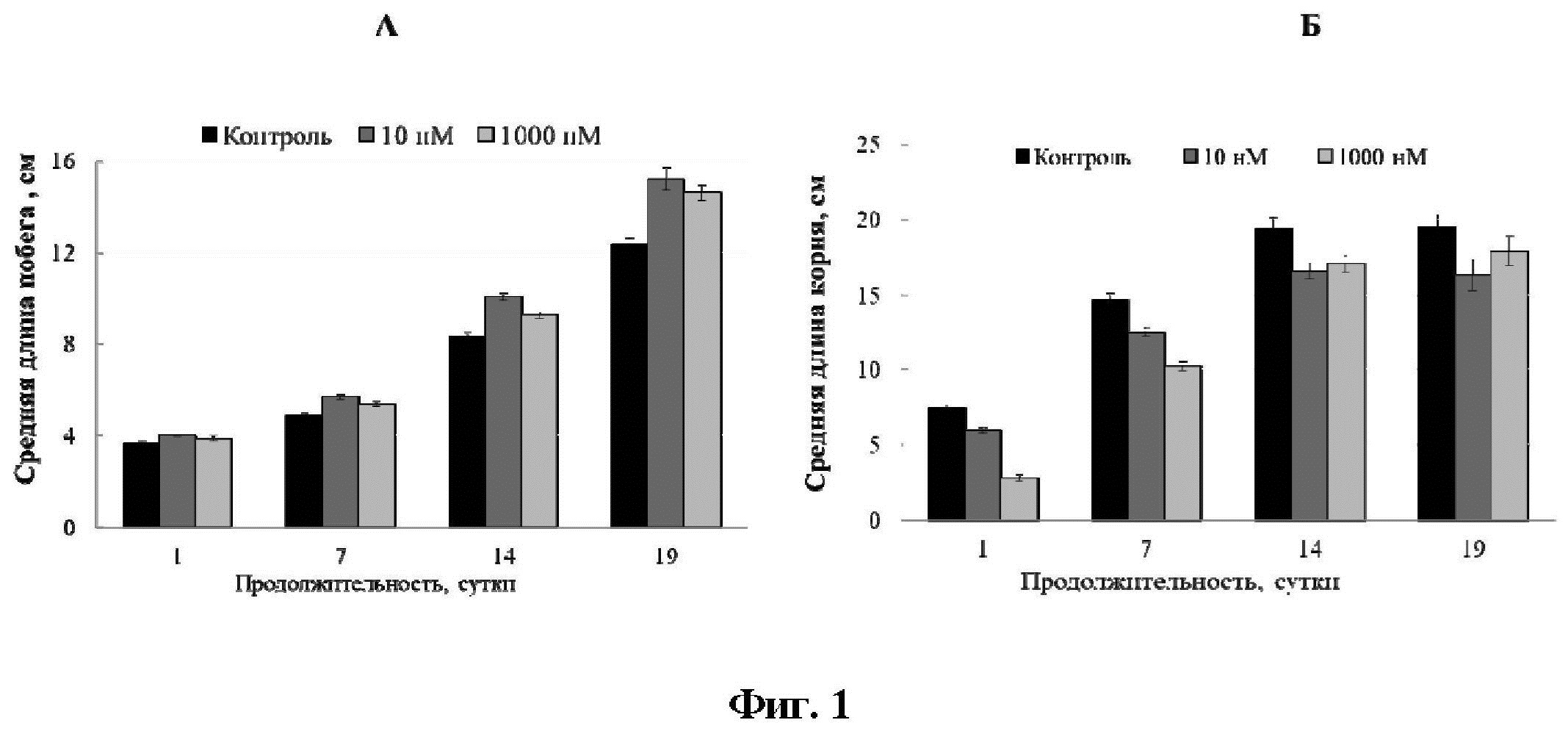
[15]На Фиг. 1 показаны гистограммы, отображающие ростовые показатели растений картофеля (А - побег, Б - корневая система) в период адаптации.
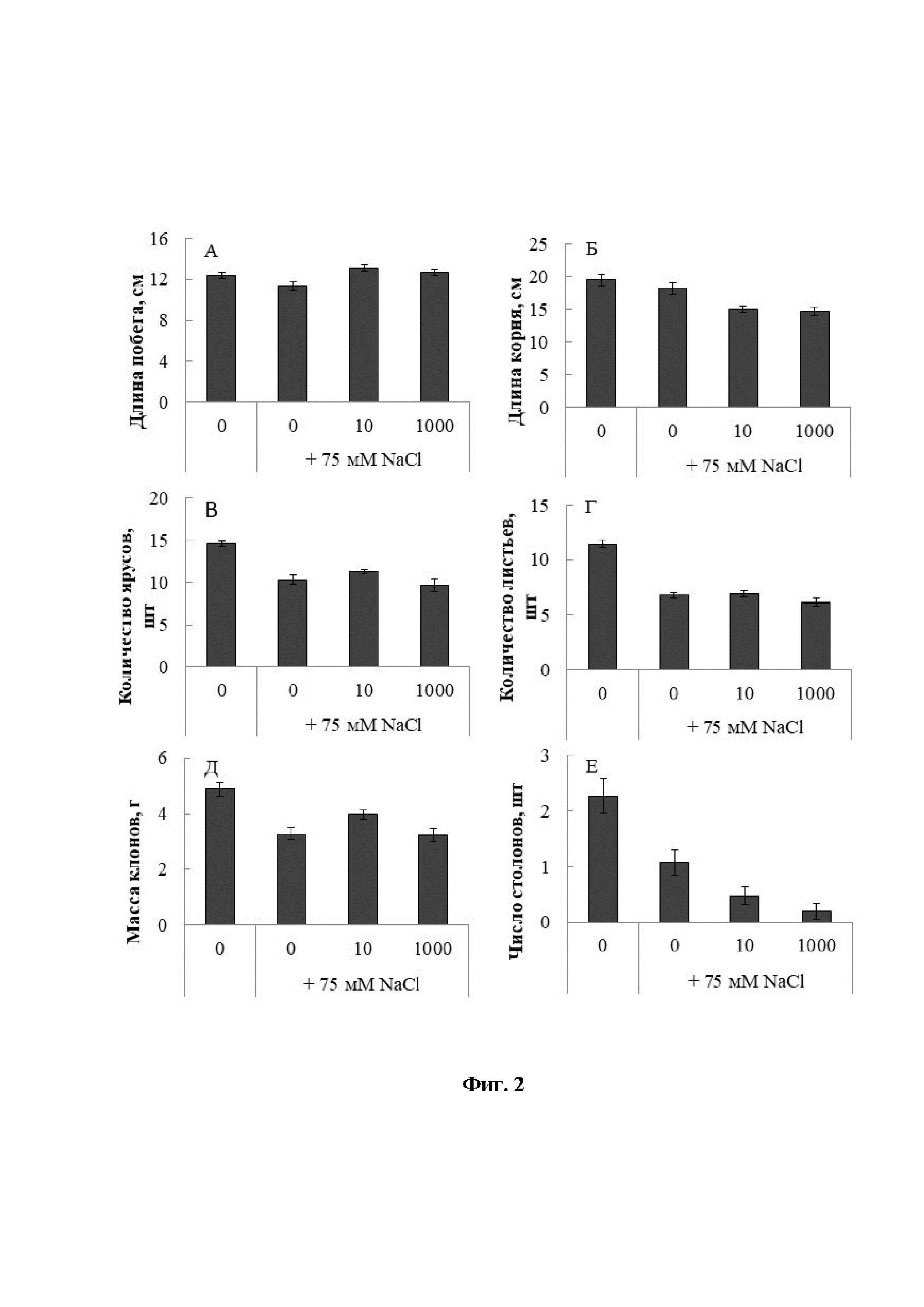
[16]На Фиг. 2 показаны гистограммы, отражающие ростовые характеристики растений картофеля (А - побег, Б - корневую систему, В - ярусы; Г - листья; Д - массу; Е - столоны).
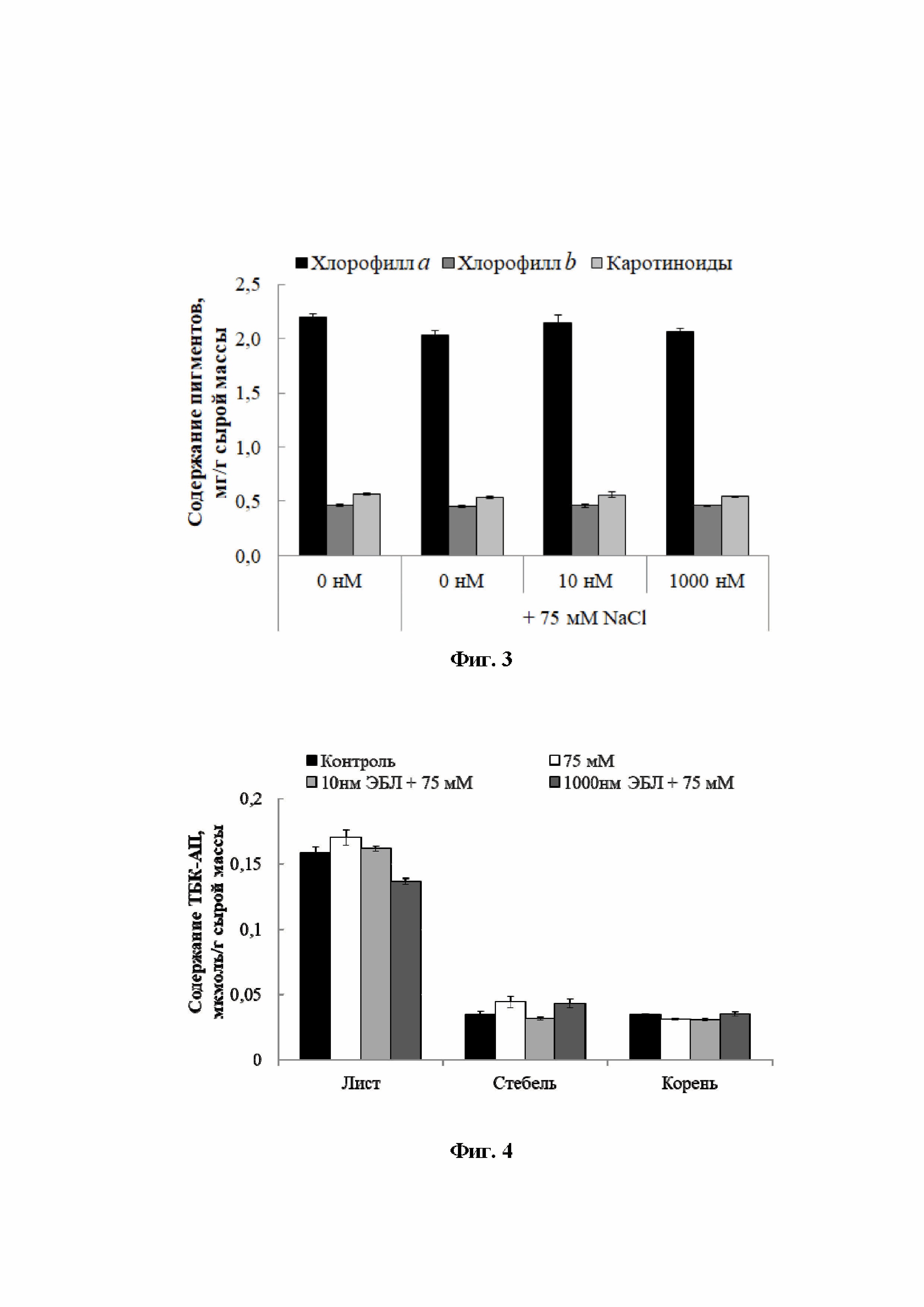
[17]На Фиг. 3 показаны гистограммы, отражающие содержание фотосинтетических пигментов (хлорофиллов , b и каротиноидов) в листьях растений картофеля.
[18]На Фиг. 4 и на Фиг. 5 показаны гистограммы, отражающие физиологическое состояние растений картофеля на примере содержания ТБК-АП (тиобарбитуровой кислоты - активных продуктов) и пролина.
[19]Примеры осуществления изобретения иллюстрируют действие на растения картофеля (ЭБЛ) разных концентраций в условиях in vitro. Проведены испытания длительного влияния 24-эпибрассинолида (10 и 1000 нМ) в условиях in vitro, при последующем солевом стрессе (75 мМ NaCl) в условиях гидропоники.
[20]Опыты проведены на растениях Soianum tuberosum L. среднеспелого сорта Луговской (идентификатор 8301891). При микроклональном размножении картофеля в культуре in vitro на начальном этапе выращивания в питательную среду Мурасиге-Скуга (75% МС) добавляли в одном варианте 10 нМ ЭБЛ, во втором - 1000 нМ ЭБЛ. Далее все растения через 21 сутки переносили на жидкую (50% МС) питательную среду без добавления гормонов для адаптации их к условиям фитокамеры на 14 суток. На следующем этапе в питательную среду вносили 75 мМ хлорида натрия для моделирования условий засоления.
[21]Ростовые характеристики (длину побега, корня) фиксировали сразу при переносе растений в условия гидропоники на этапе адаптации и через 7 или 14 суток. После воздействия хлоридом натрия дополнительно оценивали ростовые (количество ярусов и сформированных листьев, массы микроклонов, развитие столонов) и физиологические параметры (содержание пигментов, пролина, величину перекисного окисления липидов).
[22]Экстракцию фотосинтезирующих пигментов осуществляли 96% этанолом. Пробы растирали в керамических ступках с добавлением жидкого азота и переносили не менее 15 мг в пробирки Эппендорфа объемом 2 мл. В пробирки добавляли 2 мл 96% этанола для экстракции пигментов. Затем растительный материал осаждали на центрифуге MiniSpin (Eppendorf, Германия) в течение 10 минут при 8000 g. Оптическую плотность надосадочного раствора измеряли на спектрофотометре Genesys 10S UV-Vis (ThermoScientific, Германия) при длине волн: 470.0, 648.6 и 664.2 нм. В качестве контрольного образца использовали 96% этанол. Для расчета количества фотосинтетических пигментов использовали формулы, предложенные Lichtenthaler (1987).
[23]Для экстракции и определения свободного пролина навеску растительного материала (150-200 мг) растирали в жидком азоте, переносили в охлажденную пробирку Эппендорфа и добавляли 1,5 мл 3% сульфосалициловой кислоты. Далее содержимое пробирки перемешивали на вортексе и центрифугировали (15 минут, при 10000 g, 4°С). Переносили 500 мкл экстракта в стеклянные пробирки, затем добавляли 500 мкл нингидринового реактива и 500 мкл ледяной уксусной кислоты. Каждый образец был в трехкратной аналитической повторности. В контрольные образцы вместо растительного экстракта добавляли 0,5 мл 3% сульфо салициловой кислоты. Пробирки кипятили 60 минут на водяной бане под вытяжкой, затем их переносили на ледяную баню для быстрого охлаждения. Далее добавляли в каждую пробирку 1,5 мл толуола, закрывали пробирку и интенсивно перемешивается на вортексе 20 секунд. Оставляли на несколько минут до полного разделения фаз, после чего переносили верхнюю фазу с толуолом в пробирки Эппендорфа. Оптическую плотность измеряли при 520 нм на спектрофотометре Genesys 10S UV-Vis (ThermoScientific, Германия). Содержание пролина рассчитывали согласно Bates с соавторами (1973).
[24]Величину перекисного окисления липидов (ПОЛ) измеряли спектрофотометрическим методом по концентрации ТБК-АП. Ткани листа, стебля и корня растирали в керамической ступке с добавлением жидкого азота. Переносили навеску около 100 мг в пробирку Эппендорфа и добавляли 1 мл 20% трихлоруксусной кислоты (ТХУ). Перемешивали на вортексе (Vortex Microspin FV-2400), центрифугировали 15 минут при 12000 g × 4°С. В стеклянные пробирки отбирали по 500 мкл супернатанта. В контрольные образцы вместо супернатанта добавляли равный объем ТХУ В пробирки вносили 1,5 мл 0,5% тиобарбитуровой кислоты (300 мл ТХУ, 1,5 г ТБК).
[25]Реакцию, с образованием окрашенного комплекса, проводили в течение 30 минут при 98°С на водяной бане. Оптическую плотность полученных растворов измеряли на спектрофотометре (Genesys 10S UV-Vis ThermoElectron, США) при длинах волн 532 и 600 нм. Использовали формулу для расчета концентрации ТБК-АП по Buege и Aust (1978).
[26]Спустя неделю после переноса на жидкую среду, предобработка ЭБЛ увеличивала прирост длины побега в среднем на 17% и 11% соответственно при концентрациях 10 и 1000 нМ (Фиг 1А). Эффект усиливался при продолжении эксперимента, через 19 суток длина побега составляла в среднем 15 см для двух вариантов обработки гормона, в то время как у необработанных растений данный параметр был ниже, и составил 12,4 см. При этом, после прекращения гормонального воздействия, скорость роста корня была замедлена (Фиг. 1Б).
[27]Избыточное содержание хлорида натрия в среде приводит к нарушению ростовых процессов растений, отражающих их продуктивность. Наиболее выраженное действие хлорида натрия (75 мМ NaCl) было продемонстрировано на формировании ярусов и накоплении биомассы (Фиг. 2В, Г). Так, первый показатель сократился на 30%, а второй на 33% относительно контроля. ЭБЛ в концентрации 10 нМ снижал негативный эффект на накопление биомассы и формирование ярусов при отложенном действии хлорида натрия.
[28]Хлоридное засоление в меньшей степени оказывало влияние на длину осевых органов (Фиг. 2А, Б). При этом ЭБЛ при 10 нм и 1000 нм увеличивал прирост побега на 14% и 11%, соответственно, относительно солевого контроля, однако, дополнительно замедлял рост корня соответственно на 17% и 18%.
[29]Далее оценивали влияние ЭБЛ на растения при отложенном действии хлорида натрия (75 мМ) по физиологическим параметрам (содержание пигментов, пролина и степень перекисного окисления липидов).
[30]Известно, что зеленые фотосинтетические пигменты, в большей степени, подвержены действию засоления, поэтому содержание хлорофиллов используют в качестве чувствительного индикатора состояния клеточного метаболизма. При анализе влияния хлоридного засоления (75 мМ NaCl) на накопление фотосинтетических пигментов в листьях растений, обработанных 10 и 1000 нМ ЭБЛ, определяли содержание хлорофилла , хлорофилла b и каротиноидов. Нами показано, что хлоридное засоление снижало содержание хлорофилла и каротиноидов на 7%. Длительное воздействие 10 нМ ЭБЛ, способствовало поддержанию количества фотосинтетических пигментов на уровне контрольных значений (Фиг. 3). При этом хлоридное засоление не оказывало значимого эффекта на уровень хлорофилла b, как у контрольных, так и у обработанных брассиностероидом растений картофеля.
[31]Снижение содержания хлорофилла , в результате действия хлорида натрия, способствовало изменению соотношения пигментов (Таблица 1). Гормональное воздействие полностью восстанавливало данный показатель при концентрации 10 нм и частично при концентрации 1000 нМ ЭБЛ.
[32]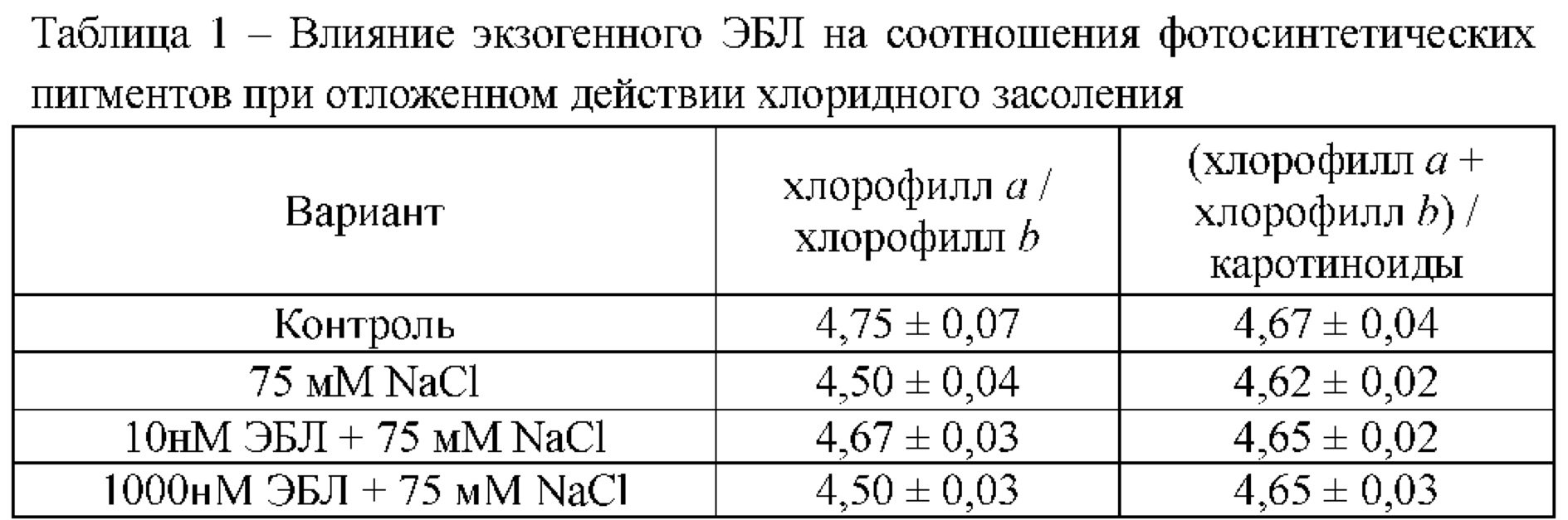
[33]Избыточные концентрации солей, поступающие в растения, стимулируют генерацию активных форм кислорода (АФК), таких как синглентный кислород, супероксидный радикал, гидроксильный радикал, перекись водорода. Названные АФК приводят к перекисному окислению белков и липидов, разрушению ДНК, окислительно-восстановительному дисбалансу. В связи с чем, далее спектрофотометрическим методом измеряли величину перекисного окисления липидов (ПОЛ) по концентрации соединений, активных в реакции с тиобарбитуровой кислотой при нагревании (ТБК-АП).
[34]При действии хлорида натрия степень ПОЛ в листьях и стебле возросла на 7% и 28% соответственно (Фиг. 4). В то время как, в корнях данный показатель, напротив, снижался на 9%. Гормональная предобработка растений способствовала снижению степени ПОЛ. Однако в зависимости от концентрации гормона эффект характеризовался органоспецифичностью. Так, в листьях и стеблях 10 нМ ЭБЛ снижал значения ПОЛ относительно солевого контроля. В то время как, 1000 нМ ЭБЛ значительно понижал содержание ТБК-АП только в листьях (Фиг. 4).
[35]Типичным ответом растений на стрессовые условия является накопление пролина - иминокислоты, которая выполняет различные функции, в том числе является осмопротектором, стабилизатором белков и ингибитором перекисного окисления липидов. В нашем исследовании было установлено, что содержание пролина повышалось в листьях в 9 раз, в стеблях и корнях в 4 раза в ответ на действие хлорида натрия, относительно контрольных значений (Фиг. 5). Гормональная обработка ЭБЛ в концентрации 10 нМ, при последующем хлоридном засолении, снижала содержание пролина в листьях, стеблях и корнях, примерно на 10% (Фиг. 5).
[36]Таким образом, экспериментально показано, что длительная прикорневая обработка ЭБЛ способствовала снижению негативного эффекта отложенного действия хлорида натрия на формирование ярусов и накопление биомассы растений картофеля. Также, в растениях предобработанных 10 нМ ЭБЛ, при хлоридном засолении сохраняется содержание фотосинтетических пигментов (хлорофилла и каротиноидов) и понижается степень перекисного окисления липидов, свидетельствующая о величине окислительного стресса. О снижении негативного воздействия хлорида натрия у обработанных ЭБЛ растений, так же указывает частичное уменьшение содержания эндогенного пролина.
[37]На основании этих результатов предложен способ повышения продуктивности растений картофеля и его агрономической устойчивости в условиях хлоридного засоления согласно заявленной формуле изобретения.
[38]Использованные источники:
[39]1. Пат. BY 022248 Фунгицидная композиция и способ ее применения Опубл.: 30.11.2015.
[40]2. Пат. RU 2728677. Сорбционно-стимулирующий препарат для предпосевной обработки семян яровой пшеницы на основе брассиностероидов Опубл.: 30.07.2020.
[41]3. Пат. RU 2349088. Способ повышения термоустойчивости овощных культур Опубл.: 20.03.2009.
[42]4. Пат. RU 2522519. Способ повышения устойчивости растений рапса к интенсивному хлоридному засолению Опубл.: 20.07.2014.
[43]5. Lichtenthaler Н.K. Chlorophylls and Carotenoids, the Pigments of Photosynthetic Biomembranes / Methods Enzymology // Eds. Douce R., Packer L. - New York: Academik. - 1987. - P. 350 - 382.
[44]6. Bates L.S. Rapid determination of free proline for water stress studies / L.S. Bates, R.P Waldran, I.D. Teare // Plant Soil. 1973. Vol. 39. P. 205 - 208.
[45]7. Microsomal lipid peroxidation / J.A. Buege, S.D. Aust // Methods Enzymol. 1978. Y 52. P. 302-310.


{kind=link}
{kind=link}
{kind=link}
{kind=link}