[1]Область техники, к которой относится изобретение
[2]Представленное изобретение относится к области медицины и биотехнологии, а именно к технологиям управления активностью возбудимых тканей, и может применяться для управления секрецией инсулина β-клетками поджелудочной железы.
[4]Свыше трехсот миллионов людей в мире болеют сахарным диабетом - комплексным заболеванием, связанным с нарушением усвоения глюкозы. Следствием заболевания является гипергликемия, приводящая к дезорганизации всех основных видов обмена веществ: углеводного, белкового, липидного и водно-солевого.
[5]В основе патогенетического механизма развития диабета первого типа лежит недостаточность синтеза и секреции инсулина эндокринными клетками поджелудочной железы (β-клетки поджелудочной железы), вызванная их разрушением в результате воздействия тех или иных факторов (вирусная инфекция, стресс, аутоиммунная агрессия и другие). Распространенность сахарного диабета первого типа достигает 10-15 % всех случаев сахарного диабета. Это заболевание характеризуется проявлением основных симптомов в детском или подростковом возрасте, быстрым развитием осложнений на фоне декомпенсации углеводного обмена. Основным методом лечения являются инъекции инсулина, нормализующие обмен веществ организма. В отсутствие лечения диабет первого типа быстро прогрессирует и приводит к возникновению тяжелых осложнений, таких как кетоацидоз и диабетическая кома.
[6]Основой патогенеза диабета второго типа является снижение чувствительности инсулинозависимых тканей к действию инсулина (инсулинорезистентность). В начальной стадии болезни инсулин синтезируется в обычных или даже повышенных количествах. Диета и снижение массы тела пациента на начальных стадиях болезни помогают нормализовать углеводный обмен, восстановить чувствительность тканей к действию инсулина и снизить синтез глюкозы на уровне печени. Однако в ходе прогрессирования заболевания биосинтез инсулина β-клетками поджелудочной железы снижается, что делает необходимым назначение заместительной гормональной терапии препаратами инсулина. Диабет второго типа составляет 85-90% всех случаев сахарного диабета у взрослого населения и наиболее часто проявляется среди лиц старше 40 лет, как правило, сопровождается ожирением. Заболевание развивается медленно, течение легкое. В клинической картине преобладают сопутствующие симптомы; кетоацидоз развивается редко. Стойкая гипергликемия с годами приводит к развитию микро- и макроангиопатии, нефро- и нейропатии, ретинопатии и других осложнений.
[7]Кроме этого, существует еще несколько видов диабета, таких как MODY-диабет и гестационный сахарный диабет, но все они клинически проявляются гипергликемией и мочеизнурением.
[8]В настоящее время лечение сахарного диабета в подавляющем большинстве случаев является симптоматическим и направлено на устранение имеющихся симптомов без устранения причины заболевания, так как эффективного лечения диабета еще не разработано. Основными задачами при лечении сахарного диабета являются: компенсация углеводного обмена, профилактика и лечение осложнений, нормализация массы тела. Компенсация углеводного обмена достигается двумя путями: путем обеспечения клеток инсулином, различными способами в зависимости от типа диабета, и путем обеспечения равномерного одинакового поступления углеводов, что достигается соблюдением диеты.
[9]В случае применения медикаментозной терапии с целью стимуляции секреции β-клетками поджелудочной железы дополнительного инсулина и восстановления нормальной концентрации глюкозы в крови используются различные пероральные сахароснижающие препараты:
[10]- Производные сульфонилмочевины (Толбутамид, Карбутамид, Хлорпропамид, Глибенкламид, Глипизид, Гликлазид, Гликвидон, Глимепирид) повышающие секрецию инсулина бета-клетками поджелудочной железы.
[11]- Прандиальные регуляторы гликемии (Репаглинид, Натеглинид) являющиеся секретагогами, обладающими быстрым всасыванием и коротким периодом сахаропонижающего действия.
[12]- Бигуаниды (метформин) снижающие абсорбцию глюкозы в кишечнике и ее производство в печени, а также повышающие чувствительность тканей к действию инсулина.
[13]- Тиазолидиндионы (росиглитазон, пиоглитазон) стимулирующие генетические механизмы, участвующие в метаболизме глюкозы, а также повышающие чувствительность тканей к глюкозе.
[14]- Ингибиторы α-гликозидазы (акарбоза) угнетающие кишечные ферменты, расщепляющие сложные углеводы до глюкозы, тем самым снижающие всасываемость глюкозы на уровне кишечника.
[15]При неэффективности данных препаратов назначается заместительная инсулинотерапия.
[16]Продукция гормона инсулина осуществляется β-клетками эндокринной части поджелудочной железы и наличие инструмента управления их активностью имеет важнейшее значение для лечения сахарного диабета. Активация клеток регулируется деполяризацией мембраны с последующим входом кальция в цитоплазму, где кальций является вторичным мессенджером в регуляции их метаболизма. Существует необходимость в методах, позволяющих управлять концентрацией кальция на внутриклеточном уровне. Однако в настоящий момент такие технологии практически отсутствуют, а имеющиеся подходы обладают недостатками, ограничивающими их широкое применение.
[17]Из уровня техники известны решения, позволяющие управлять секрецией инсулина β-клетками с применением методов оптогенетики. В статье Reinbothe с соавторами [Reinbothe TM, Safi F, Axelsson AS, Mollet IG, Rosengren AH. Optogenetic control of insulin secretion in intact pancreatic islets with β-cell-specific expression of Channelrhodopsin-2. Islets. 2014; 6(1):e28095. doi: 10.4161/isl.28095. PMID: 25483880; PMCID: PMC4593566] описан подход, основанный на оптогенетике, который позволяет проводить специфическое исследование β-клеток в островках Лангерганса. Авторы использовали трансгенных мышей, экспрессирующих светочувствительный катионный канал каналородапсин-2 (ChR2) под контролем промотора инсулина. Световая стимуляция островков трансгенных мышей ChR2 вызывала быстрое увеличение внутриклеточного кальция и усиливала секрецию инсулина в островках. β-клетки мышей, страдающих диабетом на диете с высоким содержанием жиров, демонстрировали 3,5-кратное увеличение индуцированного светом притока кальция по сравнению с мышами на контрольной диете. Кроме того, свет индуцировал повышение секреции инсулина.
[18]В исследовании Kushibiki с соавторами [Kushibiki T, Okawa S, Hirasawa T, Ishihara M. Optogenetic control of insulin secretion by pancreatic β-cells in vitro and in vivo. Gene Ther. 2015 Jul; 22(7):553-9. doi: 10.1038/gt.2015.23. Epub 2015 Mar 26. PMID: 25809465.] оценивалась способность методов оптогенетики контролировать секрецию инсулина и гомеостаз глюкозы в крови путем регулирования внутриклеточных концентраций ионов кальция в линии β-клеток поджелудочной железы мыши (MIN6), трансфицированных оптогенетическим канальным белком ChR2. Клетки MIN6, трансфицированные ChR2 (ChR2-MIN6), секретировали инсулин после облучения лазером (470 нм). Увеличение кальция сопровождалось повышенным уровнем матричных РНК, которые кодируют кальций/кальмодулинзависимую протеинкиназу II дельта и аденилатциклазу 1. Клетки ChR2-MIN6, суспендированные в матригеле, трансплантировали мышам с диабетом, индуцированным стрептозотоцином. которые затем подвергались тесту на толерантность к глюкозе. Лазерное облучение этих мышей вызывало значительное снижение уровня глюкозы в крови, а облученные имплантированные клетки экспрессировали инсулин.
[19]Результаты демонстрируют полезность и универсальность оптогенетики для изучения механизмов гомеостаза глюкозы и для разработки методов лечения метаболических заболеваний, таких как сахарный диабет. Существенным недостатком данных подходов является использование каналородопсинов, не встречающихся у млекопитающих и вызывающих иммунную реакцию, которая в относительно короткий срок приводит к гибели клеток, экспрессирующих каналородопсины. Перспективным подходом для решения данной проблемы может являться применение термогенетических кальциевых переключателей.
[20]Термогенетика представляет собой подход для регуляции активности клеток с помощью термоактивируемых каналов семейства TRP (Transient Receptor Potential), применяемый как для модельных клеточных культур, так и для животных in vivo. TRP являются неселективными катионными каналами, способными при активации пропускать ионы кальция, натрия, калия и магния. Применение TRP каналов, позволяющих управлять внутриклеточной концентрацией кальция, является перспективным подходом для контроля активности клеток, в частности, секрецией инсулина β-клетками.
[21]Из уровня техники известны подходы, позволяющие управлять клетками при помощи термочувствительных ионных каналов, однако данные решения не используются для контроля активности β-клеток поджелудочной железы. Например, запатентованы способы, позволяющие стимулировать иммунные клетки с помощью экспрессии в них механо- и термочувствительных ионных каналов c последующей активацией при помощи ультразвука [патент WO2018098315A1].
[22]Одним из представленных в научной литературе подходов, наиболее близким к заявленному нами, является неинвазивная нейромодуляция глубоких отделов мозга [Yang Y, Pacia CP, Ye D, Zhu L, Baek H, Yue Y, Yuan J, Miller MJ, Cui J, Culver JP, Bruchas MR, Chen H. Sonothermogenetics for noninvasive and cell-type specific deep brain neuromodulation. Brain Stimul. 2021 Jul-Aug; 14(4):790-800. doi: 10.1016/j.brs.2021.04.021]. В данном подходе используются термочувствительные ионные каналы TRPV1 и нагрев клеток мозга при помощи высокоинтенсивного сфокусированного ультразвука. Однако, данный подход не применялся для управления активностью β-клеток поджелудочной железы.
[23]Управление клеточной активностью с использованием термоактивируемых каналов суперсемейства TRP является перспективным подходом, поскольку эти каналы присутствуют в организме человека, и их экспрессия не вызывает иммунного ответа. Необходимое для активации термочувствительных каналов излучение невидимо для живых организмов, что снижает уровень стресса животного в процессе стимуляции. За счет низкого поглощения собственными пигментами, инфракрасное и СВЧ излучение не вызывает фототоксических эффектов в верхних слоях тканей как в случае использования синего света для активации ChRs.
[24]Раскрытие изобретения
[25]Техническая проблема, решаемая настоящим изобретением, заключается в отсутствии возможности неинвазивногоуправления секрецией инсулина β-клетками поджелудочной железы.
[26]При решении заявленной технической проблемы достигается следующие технические результаты: возможность неинвазивного управления секрецией инсулина β-клетками отдельных островков Лангерганса поджелудочной железы при помощи нагрева.
[27]Для решения заявленной технической проблемы и достижения заявленного технического результата предлагается способ управления секрецией инсулина β-клетками островков Лангерганса поджелудочной железы с использованием подходов термогенетики, включающие в себя следующие действия:
[28]- создание генетических конструкций, содержащих в своем составе специфический для β-клеток промотер и гены термически активируемых каналов суперсемейства TRP;
[29]- доставку указанных конструкций с помощью аденоассоциированных вирусов в β-клетки островков Лангерганса для последующей экспрессии соответствующего канала;
[30]- нагрев β-клеток выделенных островков Лангерганса поджелудочной железы при помощи импульсов заданной частоты или в постоянном режиме, с помощью инфракрасного (ИК) лазера или сфокусированного ультразвука.
[31]Краткое описание чертежей
[32]Изобретение поясняется фигурами, где представлены:
[33]Фиг. 1. Карта генетической конструкции на основе плазмиды AAV, содержащей специфичный для β-клеток промотор Ins2, а также последовательность человеческого термически активируемого канала TRPV1 с сигнальным пептидом FLAG-tag.
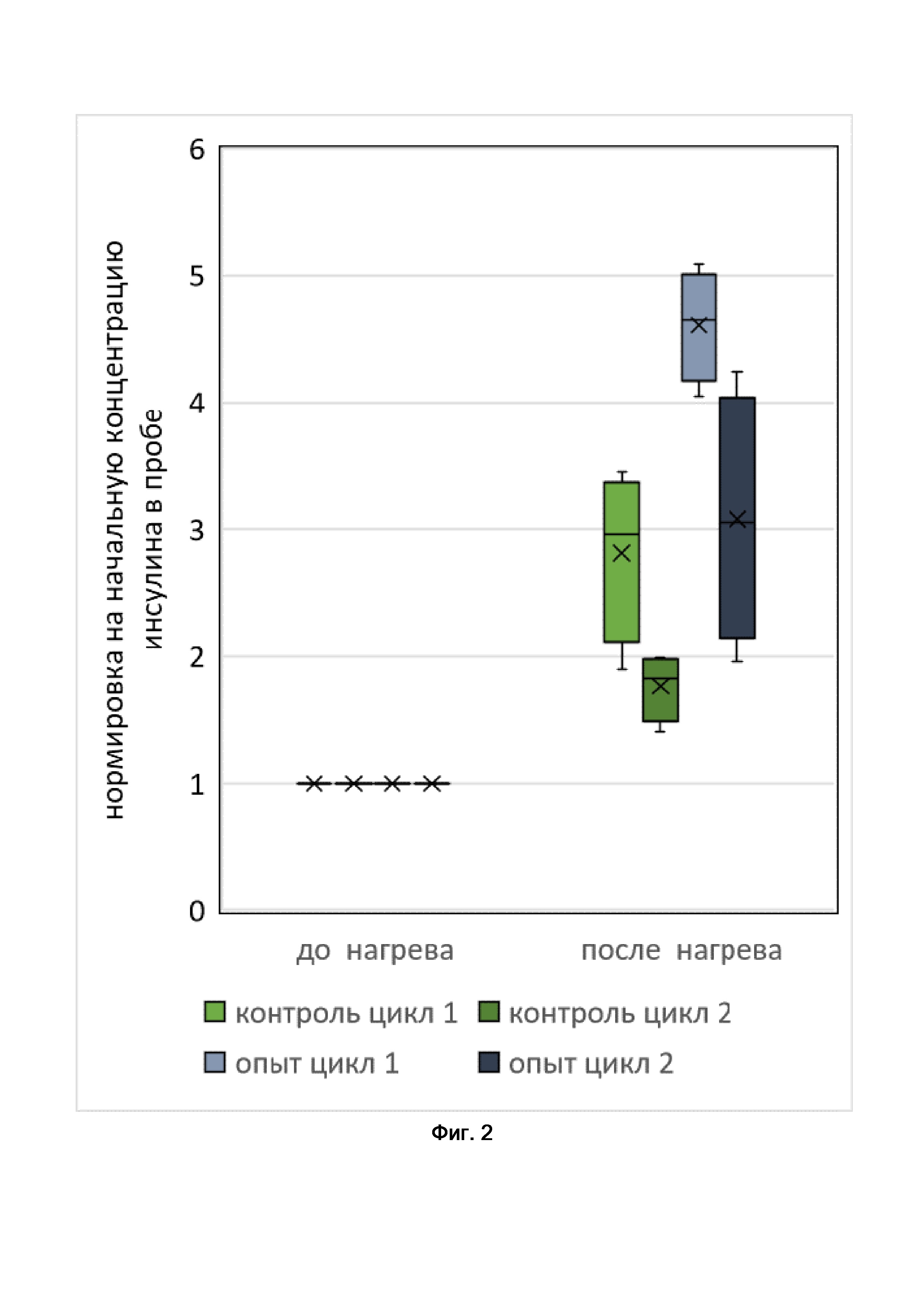
[34]Фиг. 2. Результаты нагрева отдельных островков Лангерганса трансдуцированных конструкцией AAV-Ins2-hTRPV1-FLAG. Нагрев осуществляется в режиме цикл1 (20 минут 37C ,(1 минута 43C, 1 минута 37C) х10), цикл 2 (20 минут 37C ,(10 секунд 43C, 50 секунд 37C) х20.
[35]Осуществление изобретения
[36]В общем виде способ управления секрецией инсулина β-клетками островков Лангерганса поджелудочной железы с использованием подходов термогенетики, включает следующие этапы:
[37]- создание генетической конструкции, с помощью клонирования в коммерчески доступный вектор специфичного для β-клеток промотера, гена термически активируемого канала, а также последовательности сигнального пептида FLAG, необходимой для идентификации TRPV1 при помощи иммуногистохимии;
[38]- наработку полученной генетической конструкции при помощи трансформации бактерий E. coli с последующим их наращиванием и выделением плазмидной ДНК;
[39]- генерацию вирусного вектора, подходящего для доставки генетической конструкции в β-клетки;
[40]- доставку генетической конструкции посредством трансдукции β-клеток островков Лангерганса;
[41]- бесконтактное термическое воздействие на выделенные островки Лангерганса при помощи ИК лазера или высокоинтенсивного сфокусированного ультразвука с контролем концентрации инсулина.
[43]Пример 1. Управление секрецией инсулина β-клетками островков Лангерганса при помощи термогенетики.
[44]Создание и наработка генетической конструкции, генерация вирусного вектора
[45]Для создания генетических конструктов пользовались методом AQUA cloning (Beyer et al., 2015). В качестве основы для клонирования была взята плазмида AAV, в которую были клонированы специфичный для β-клеток промотер Ins2, последовательность человеческого термически активируемого канала TRPV1, а также последовательность сигнального пептида FLAG, необходимого для идентификации TRPV1 при помощи иммуногистохимии. Карта полученной конструкции AAV-Ins2-hTRPV1-FLAG приведена на фиг. 1. Аденоассоциированные вирусы серотипа PHP.S, несущие конструкцию AAV-Ins2-hTRPV1-FLAG, были сгенерированы при помощи коммерчески доступного сервиса.
[46]Выделение и трансдукция островков Лангерганса
[47]Выделения островков Лангерганса проводили на мышах в возрасте от 2 до 3 месяцев с использованием модифицированной версии метода опубликованного Stull с соавторами [Stull ND, Breite A, McCarthy R, Tersey SA, Mirmira RG. Mouse islet of Langerhans isolation using a combination of purified collagenase and neutral protease. J Vis Exp. 2012 Sep 7; (67):4137. doi: 10.3791/4137. PMID: 22987198; PMCID: PMC3490250].
[48]Выделение проводили по следующему протоколу:
[49]На первом этапе мышь подвергалась эвтаназии посредством помещения в камеру с высоким содержанием углекислого газа. Для получения доступа к поджелудочной железе под операционным микроскопом выполнялся V образный разрез кожи и брюшной стенки. Во избежание излишнего попадания крови в изъятую поджелудочную железу производился разрез нижней полой вены и отбор крови. Далее зажимом перекрывался общий печеночный проток. В районе впадения поджелудочного протока в двенадцатиперстную кишку под протоком с помощью пинцета производили разрыв соединительных тканей и в полученное отверстие продевали нить, делали петлю. С помощью маленьких очень острых ножниц разрезали место впадения поджелудочного протока в двенадцатиперстную кишку. В полученное отверстие вводили иглу 31G на 0.5-1 см и затягивали петлю. К игле присоединялась трубка со шприцом, наполненным холодным раствором коллагеназы XI (Sigma , C7657, США) объемом 2 мл концентрации 1,5 мг/мл. Далее поджелудочная железа извлекалась и помещалась в 4 мл раствора коллагеназы, инкубировалась на водяной бане при +37С в течение 15 минут. После инкубации к поджелудочной железе добавляли раствор Кребса-Рингера бикарбонатный с добавлением HEPES (KRBH) с 0,3% бычьего сывороточного альбумина (БСА) и интенсивно трясли 15 раз для диссоциации поджелудочной железы. Полученную суспензию центрифугировали 1 минуту при 400 G, супернатант сливали, к осадку добавляли 20 мл KRBH с 0,3% БСА. Два-три раза интенсивно разбивали осадок пипетированием серологической пипеткой. Затем клеточная суспензия пропускалась через сетку с размером ячеек 0,5 мм, центрифугировали 1 минуту при 400 G. Далее проводилось разделение с помощью градиента плотности, полученный осадок ресуспезировали в 10 мл раствора для выделения лимфоцитов плотностью 1,077 г/мл (Capricorn scientific, Германия). Сверху наслаивали KRBH и центрифугировали 20 минут при 900 G. На границе растворов собирали островки Лангерганса с помощью серологической пипетки и пропускали через фильтр с ячейками 40 мкм. Отмывали островки Лангерганса с помощью среды RPMI 1640 с добавлением антибиотика, глутамина и 10 % фетальной бычьей сыворотки в чашку петри диаметром 60 мм. На заключительном этапе островки Лангерганса с нормальной физиологией отбирались вручную и переносились по 25-30 штук в одну лунку 12 луночного планшета без покрытия поверхности для дальнейшей культивации.
[50]Трансдукцию аденоассоциированным вирусом в концентрации 5,2*10^13 вирусных геномов/мл, 1,5-2 мкл раствора вируса на 0,5 мл среды и 25-30 островков Лангерганса, несущим конструкцию AAV-Ins2-hTRPV1-FLAG, проводили сразу после выделения. Изолированные островки культивировали при 37°С, 5% CO2 в течение 3-5 дней.
[51]Бесконтактное термическое воздействие на β-клетки отдельных островков Лангерганса при помощи нагрева.
[52]Для управления секрецией инсулина β-клетками трансдуцированные островки Лангерганса вирусом, несущим конструкцию AAV-Ins2-hTRPV1-FLAG, переносили в ПЦР пробирки для нагрева в ПЦР машине. Нагрев осуществляется в режиме цикл 1 (20 минут 37C , (1 минута 43C, 1 минута 37C) х10), цикл 2 (20 минут 37C ,(10 секунд 43C, 50 секунд 37C) х20. Измерения концентрации секретированного инсулина проводилось с помощью ИФА набора (Mercodia, Швеция).
[53]В проведенном эксперименте продемонстрировано, что нагрев отдельных островков Лангерганса приводит увеличению к секреции инсулина как минимум в 1,8 раза.
[55]<?xml version="1.0" encoding="UTF-8"?>
[56]<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
[57]1.3//EN" "ST26SequenceListing_V1_3.dtd">
[58]<ST26SequenceListing originalFreeTextLanguageCode="ru"
[59]dtdVersion="V1_3" fileName="Способ управления секрецией инсулина
[60]β-клетками поджелудочной железы при помощи термогенетики..xml"
[61]softwareName="WIPO Sequence" softwareVersion="2.2.0"
[62]productionDate="2023-02-13">
[63] <ApplicationIdentification>
[64] <IPOfficeCode>RU</IPOfficeCode>
[65] <ApplicationNumberText>2022125718/20(056035)</ApplicationNumberText>
[66] <FilingDate>2022-10-01</FilingDate>
[67] </ApplicationIdentification>
[68] <ApplicantFileReference>2022125718/20(056035)</ApplicantFileReference
[70] <EarliestPriorityApplicationIdentification>
[71] <IPOfficeCode>RU</IPOfficeCode>
[72] <ApplicationNumberText>2022125718/20(056035)</ApplicationNumberText>
[73] <FilingDate>2022-10-01</FilingDate>
[74] </EarliestPriorityApplicationIdentification>
[75] <ApplicantName languageCode="ru">Институт биоорганической химии им.
[76]ак. М.М.Шемякина и Ю.А.Овчинникова Российской Академии
[78] <ApplicantNameLatin>Shemyakin-Ovchinnikov Institute of Bioorganic
[79]Chemistry of the Russian Academy of Sciences</ApplicantNameLatin>
[80] <InventionTitle languageCode="ru">Способ управления секрецией
[81]инсулина β-клетками поджелудочной железы при помощи
[82]термогенетики.</InventionTitle>
[83] <SequenceTotalQuantity>1</SequenceTotalQuantity>
[84] <SequenceData sequenceIDNumber="1">
[86] <INSDSeq_length>6518</INSDSeq_length>
[87] <INSDSeq_moltype>DNA</INSDSeq_moltype>
[88] <INSDSeq_division>PAT</INSDSeq_division>
[89] <INSDSeq_feature-table>
[91] <INSDFeature_key>source</INSDFeature_key>
[92] <INSDFeature_location>1..6518</INSDFeature_location>
[95] <INSDQualifier_name>mol_type</INSDQualifier_name>
[96] <INSDQualifier_value>other DNA</INSDQualifier_value>
[98] <INSDQualifier id="q4">
[99] <INSDQualifier_name>organism</INSDQualifier_name>
[100] <INSDQualifier_value>synthetic construct</INSDQualifier_value>
[102] </INSDFeature_quals>
[104] </INSDSeq_feature-table>
[105] <INSDSeq_sequence>cctgcaggcagctgcgcgctcgctcgctcactgaggccgcccgggcaaa
[106]gcccgggcgtcgggcgacctttggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggcc
[107]aactccatcactaggggttcctgcggccgcacgcgtcggaccattaagtgccttgctgcctgaattctgc
[108]tttccttctacctctgagagagagctggggactcggctgagttaagaacccagctatcaattggaactgt
[109]gaaacagtccaagggacaaagatactaggtccccaactgcaacttcctggggaatgatgtggaaaaatgc
[110]tcagccaaggacaaagaaagcatcacccactctggaacaatgtcccctgctgtgaactggttcatcaggc
[111]catcagggccccttgttaagactctaattaccctaggactaagtagaggtgttgacgtccaatgagcgct
[112]ttctgcagacctagcaccagggaagtgtttggaaactgcagcttcagcccctctggccatctgctgacct
[113]accccacctggagcccttaatgggtcaaacagcaaagtccagggggcagagaggaggtgctttggtctat
[114]aaaggtagtggggacccagtaaccaccagccctaagtgatccaggatccgccaccatgaagaaatggagc
[115]agcacagacttgggggcagctgcggacccactccaaaaggacacctgcccagaccccctggatggagacc
[116]ctaactccaggccacctccagccaagccccagctctccacggccaagagccgcacccggctctttgggaa
[117]gggtgactcggaggaggctttcccggtggattgccctcacgaggaaggtgagctggactcctgcccgacc
[118]atcacagtcagccctgttatcaccatccagaggccaggagacggccccaccggtgccaggctgctgtccc
[119]aggactctgtcgccgccagcaccgagaagaccctcaggctctatgatcgcaggagtatctttgaagccgt
[120]tgctcagaataactgccaggatctggagagcctgctgctcttcctgcagaagagcaagaagcacctcaca
[121]gacaacgagttcaaagaccctgagacagggaagacctgtctgctgaaagccatgctcaacctgcatgacg
[122]gacagaacaccaccatccccctgctcctggagatcgcgcggcaaacggacagcctgaaggagcttgtcaa
[123]cgccagctacacggacagctactacaagggccagacagcactgcacatcgccatcgagagacgcaacatg
[124]gccctggtgaccctcctggtggagaacggagcagacgtccaggctgcggcccatggggacttctttaaga
[125]aaaccaaagggcggcctggattctacttcggtgaactgcccctgtccctggccgcgtgcaccaaccagct
[126]gggcatcgtgaagttcctgctgcagaactcctggcagacggccgacatcagcgccagggactcggtgggc
[127]aacacggtgctgcacgccctggtggaggtggccgacaacacggccgacaacacgaagtttgtgacgagca
[128]tgtacaatgagattctgatcctgggggccaaactgcacccgacgctgaagctggaggagctcaccaacaa
[129]gaagggaatgacgccgctggctctggcagctgggaccgggaagatcggggtcttggcctatattctccag
[130]cgggagatccaggagcccgagtgcaggcacctgtccaggaagttcaccgagtgggcctacgggcccgtgc
[131]actcctcgctgtacgacctgtcctgcatcgacacctgcgagaagaactcggtgctggaggtgatcgccta
[132]cagcagcagcgagacccctaatcgccacgacatgctcttggtggagccgctgaaccgactcctgcaggac
[133]aagtgggacagattcgtcaagcgcatcttctacttcaacttcctggtctactgcctgtacatgatcatct
[134]tcaccatggctgcctactacaggcccgtggatggcttgcctccctttaagatggaaaaaattggagacta
[135]tttccgagttactggagagatcctgtctgtgttaggaggagtctacttctttttccgagggattcagtat
[136]ttcctgcagaggcggccgtcgatgaagaccctgtttgtggacagctacagtgagatgcttttctttctgc
[137]agtcactgttcatgctggccaccgtggtgctgtacttcagccacctcaaggagtatgtggcttccatggt
[138]attctccctggccttgggctggaccaacatgctctactacacccgcggtttccagcagatgggcatctat
[139]gccgtcatgatagagaagatgatcctgagagacctgtgccgtttcatgtttgtctacatcgtcttcttgt
[140]tcgggttttccacagcggtggtgacgctgattgaagacgggaagaatgactccctgccgtctgagtccac
[141]gtcgcacaggtggcgggggcctgcctgcaggccccccgatagctcctacaacagcctgtactccacctgc
[142]ctggagctgttcaagttcaccatcggcatgggcgacctggagttcactgagaactatgacttcaaggctg
[143]tcttcatcatcctgctgctggcctatgtaattctcacctacatcctcctgctcaacatgctcatcgccct
[144]catgggtgagactgtcaacaagatcgcacaggagagcaagaacatctggaagctgcagagagccatcacc
[145]atcctggacacggagaagagcttccttaagtgcatgaggaaggccttccgctcaggcaagctgctgcagg
[146]tggggtacacacctgatggcaaggacgactaccggtggtgcttcagggtggacgaggtgaactggaccac
[147]ctggaacaccaacgtgggcatcatcaacgaagacccgggcaactgtgagggcgtcaagcgcaccctgagc
[148]ttctccctgcggtcaagcagagtttcaggcagacactggaagaactttgccctggtcccccttttaagag
[149]aggcaagtgctcgagataggcagtctgctcagcccgaggaagtttatctgcgacagttttcagggtctct
[150]gaagccagaggacgctgaggtcttcaagagtcctgccgcttccggggagaaggattacaaggacgacgac
[151]gataaggattacaaggacgacgacgataaggattacaaggacgacgacgataagtaaagatctacgggtg
[152]gcatccctgtgacccctccccagtgcctctcctggccctggaagttgccactccagtgcccaccagcctt
[153]gtcctaataaaattaagttgcatcattttgtctgactaggtgtccttctataatattatggggtggaggg
[154]gggtggtatggagcaaggggcaagttgggaagacaacctgtagggcctgcggggtctattgggaaccaag
[155]ctggagtgcagtggcacaatcttggctcactgcaatctccgcctcctgggttcaagcgattctcctgcct
[156]cagcctcccgagttgttgggattccaggcatgcatgaccaggctcagctaatttttgtttttttggtaga
[157]gacggggtttcaccatattggccaggctggtctccaactcctaatctcaggtgatctacccaccttggcc
[158]tcccaaattgctgggattacaggcgtgaaccactgctcccttccctgtccttctgattttgtaggtaacc
[159]acgtgcggaccgagcggccgcaggaacccctagtgatggagttggccactccctctctgcgcgctcgctc
[160]gctcactgaggccgggcgaccaaaggtcgcccgacgcccgggctttgcccgggcggcctcagtgagcgag
[161]cgagcgcgcagctgcctgcaggggcgcctgatgcggtattttctccttacgcatctgtgcggtatttcac
[162]accgcatacgtcaaagcaaccatagtacgcgccctgtagcggcgcattaagcgcggcgggtgtggtggtt
[163]acgcgcagcgtgaccgctacacttgccagcgccctagcgcccgctcctttcgctttcttcccttcctttc
[164]tcgccacgttcgccggctttccccgtcaagctctaaatcgggggctccctttagggttccgatttagtgc
[165]tttacggcacctcgaccccaaaaaacttgatttgggtgatggttcacgtagtgggccatcgccctgatag
[166]acggtttttcgccctttgacgttggagtccacgttctttaatagtggactcttgttccaaactggaacaa
[167]cactcaaccctatctcgggctattcttttgatttataagggattttgccgatttcggcctattggttaaa
[168]aaatgagctgatttaacaaaaatttaacgcgaattttaacaaaatattaacgtttacaattttatggtgc
[169]actctcagtacaatctgctctgatgccgcatagttaagccagccccgacacccgccaacacccgctgacg
[170]cgccctgacgggcttgtctgctcccggcatccgcttacagacaagctgtgaccgtctccgggagctgcat
[171]gtgtcagaggttttcaccgtcatcaccgaaacgcgcgagacgaaagggcctcgtgatacgcctattttta
[172]taggttaatgtcatgataataatggtttcttagacgtcaggtggcacttttcggggaaatgtgcgcggaa
[173]cccctatttgtttatttttctaaatacattcaaatatgtatccgctcatgagacaataaccctgataaat
[174]gcttcaataatattgaaaaaggaagagtatgagtattcaacatttccgtgtcgcccttattccctttttt
[175]gcggcattttgccttcctgtttttgctcacccagaaacgctggtgaaagtaaaagatgctgaagatcagt
[176]tgggtgcacgagtgggttacatcgaactggatctcaacagcggtaagatccttgagagttttcgccccga
[177]agaacgttttccaatgatgagcacttttaaagttctgctatgtggcgcggtattatcccgtattgacgcc
[178]gggcaagagcaactcggtcgccgcatacactattctcagaatgacttggttgagtactcaccagtcacag
[179]aaaagcatcttacggatggcatgacagtaagagaattatgcagtgctgccataaccatgagtgataacac
[180]tgcggccaacttacttctgacaacgatcggaggaccgaaggagctaaccgcttttttgcacaacatgggg
[181]gatcatgtaactcgccttgatcgttgggaaccggagctgaatgaagccataccaaacgacgagcgtgaca
[182]ccacgatgcctgtagcaatggcaacaacgttgcgcaaactattaactggcgaactacttactctagcttc
[183]ccggcaacaattaatagactggatggaggcggataaagttgcaggaccacttctgcgctcggcccttccg
[184]gctggctggtttattgctgataaatctggagccggtgagcgtgggtctcgcggtatcattgcagcactgg
[185]ggccagatggtaagccctcccgtatcgtagttatctacacgacggggagtcaggcaactatggatgaacg
[186]aaatagacagatcgctgagataggtgcctcactgattaagcattggtaactgtcagaccaagtttactca
[187]tatatactttagattgatttaaaacttcatttttaatttaaaaggatctaggtgaagatcctttttgata
[188]atctcatgaccaaaatcccttaacgtgagttttcgttccactgagcgtcagaccccgtagaaaagatcaa
[189]aggatcttcttgagatcctttttttctgcgcgtaatctgctgcttgcaaacaaaaaaaccaccgctacca
[190]gcggtggtttgtttgccggatcaagagctaccaactctttttccgaaggtaactggcttcagcagagcgc
[191]agataccaaatactgtccttctagtgtagccgtagttaggccaccacttcaagaactctgtagcaccgcc
[192]tacatacctcgctctgctaatcctgttaccagtggctgctgccagtggcgataagtcgtgtcttaccggg
[193]ttggactcaagacgatagttaccggataaggcgcagcggtcgggctgaacggggggttcgtgcacacagc
[194]ccagcttggagcgaacgacctacaccgaactgagatacctacagcgtgagctatgagaaagcgccacgct
[195]tcccgaagggagaaaggcggacaggtatccggtaagcggcagggtcggaacaggagagcgcacgagggag
[196]cttccagggggaaacgcctggtatctttatagtcctgtcgggtttcgccacctctgacttgagcgtcgat
[197]ttttgtgatgctcgtcaggggggcggagcctatggaaaaacgccagcaacgcggcctttttacggttcct
[198]ggccttttgctggccttttgctcacatgt</INSDSeq_sequence>
[201]</ST26SequenceListing>


{kind=link}
{kind=link}