Изобретение относится к области молекулярной генетики и может быть использовано в профилактической медицине при создании вакцин. В качестве объекта, направленное воздействие на который приводит к утрате способности патогенного микроорганизма проявлять вирулентные свойства, выбран ген mutR. Он может быть использован в качестве мишени для разработки и применения селективных ингибиторов с целью профилактики или лечения стрептококковых заболеваний.
На основе полученных в результате инактивации гена mutR авирулентных штаммов Streptococcus pyogenes реально получение живой стрептококковой вакцины для профилактики заболеваний, вызываемых данным микроорганизмом.
Стабилизированное снижение или полная потеря вирулентности служит основой для применения штаммов в качестве вакцин. Таким образом, авирулентные штаммы активно используются в качестве живых вакцин.
Живые вакцины создают напряженный иммунитет, сходный с естественным постинфекционным. Как правило, живые вакцины вводят однократно, т.к. вакцинный штамм персистирует в организме. Положительные стороны таких вакцин: по механизму действия на организм авирулентный штамм напоминает "дикий" штамм; авирулентный штамм может приживляться в организме, в результате организм длительно сохраняет иммунитет. Используются небольшие дозы для вакцинации (обычно ее проводят однократно), и поэтому вакцинацию легко проводить организационно. Последнее позволяет рекомендовать данный тип вакцин для дальнейшего использования.
Известна живая вакцина для профилактики заболеваний, вызываемых менингококками. В этом случае используется авирулентный штамм, мутантный по гену phoP. PhoP входит в состав двухкомпонентной системы регуляции транскрипции и является важным регулятором экспрессии генов вирулентности у различных штаммов менингококков. Авирулентный штамм, мутантный по гену phoP, был получен методом замены аллелей [Newcombe J., Eales-Reynolds L.J., Wootton L., Gorringe A.R., Funnell S.G., Taylor S.C., McFadden J.J. Infection with an avirulent phoP mutant of Neisseria meningitidis confers broad cross-reactive immunity // Infect. Immun. - 2004. - 72. P.338-344].
Streptococcus pyogenes (β - гемолитический стрептококк серогруппы A) - один из наиболее часто встречающихся возбудителей бактериальных инфекций человека. S. pyogenes вызывает такие распространенные заболевания, как тонзилло-фарингит, скарлатина, импетиго, рожистое воспаление, флегмоны, артрит и др. В частности, в США ежегодно у 1,8 млн. чел. диагностируют стрептококковый фарингит с социально-экономическим ущербом более 100 млн. долларов [Neuner J.M., Hamel M.B., Phillips R.S., Bona K., Aronson M.D. Diagnosis and management of adults with pharyngitis. A cost-effectiveness analysis // Ann. Intern. Med. - 2003. - 139. P.113-122]. К осложнениям инфекций, вызываемых S. pyogenes, относят острую ревматическую лихорадку, гломерулонефрит и хорею. В последние годы отмечен рост заболеваемости тяжелыми инвазивными стрептококковыми инфекциями (некротизирующий фасцит, синдром токсического шока). Так, например, 700 тыс. чел. ежегодно страдают от инвазивных форм стрептококковых заболеваний, и при этом смертность достигает 50% [Davies H.D., Mcgeer A., Schwartz B. Invasive group a streptococcal infections in Ontario, Canada. Ontario group a streptococcal study group // N. Engl. J. Med. - 1996. - 335. - P.547-554; Carapetis J.R., Steer A.C., Mulholland E.K., Weber M. The global burden of group a streptococcal diseases // Lancet. Infect. Dis. - 2005. - 5. P.685-694]. Все вышеперечисленное повышает интерес к инфекциям, вызываемым S. pyogenes.
Борьба со стрептококковыми инфекциями ведется медикаментозно. Для лечения применяют антибактериальную терапию, используются антибиотики, такие как пенициллины, эритромицин и линкосамиды (линкомицин, клиндамицин) [Choby B.A. Diagnosis and treatment of streptococcal pharyngitis // Am Fam Physician. - 2009. - 79. - P.383-90]. Важным является не допускать развитие заболевания и предотвращать появление осложнений, чему помогает применение живых вакцин, созданных на основе авирулентных штаммов.
Во взаимодействие S. pyogenes с организмом человека вовлечено большое количество факторов патогенности, которые способствуют адгезии и колонизации микроорганизма, генерализации инфекции, уклонению от иммунного ответа и длительной персистенции патогена в организме человека [Cunningham M.W. Pathogenesis of group A streptococcal infections // Clin. Microbiol. Rev. - 2000. - 13. - P.470-511].
Экспрессия этих факторов патогенности контролируется регуляторными молекулами или комплексами молекул, которые действуют в ответ на изменение условий обитания микроорганизма. В частности, для регуляции транскрипции генов патогенности S. pyogenes использует 13 двухкомпонентных регуляторных систем и более 40 белков-регуляторов, в том числе глобальные белки-регуляторы Mga, Rgg и белки семейства RALP.
По данным компьютерного моделирования перечисленные белки содержат участок "спираль-поворот-спираль", за счет которого они взаимодействуют с промоторными областями отдельных генов, в том числе генов патогенности, влияя на уровни их транскрипции и, как следствие, на уровни экспрессии кодируемых ими белков.
Белок-регулятор Mga S. pyogenes контролирует экспрессию поверхностных и секретируемых белков, участвующих в колонизации тканей человека, а также белков, позволяющих избегать воздействия иммунной системы. Mga контролирует экспрессию основного антифагоцитарного белка S. pyogenes (М-белок, ген emm), коллаген-подобного белка (ген scll/sclA) и фактора опалесценции (ген sof). Кроме того, Mga регулирует экспрессию М-подобных белков, способных связывать иммуноглобулины A и G человека (гены mrp, arp и enn), C5а пептидазы (ген scpA), ингибитора комплемента (ген sic) и других [McIver K.S., Scott J.R. Role of mga in growth phase regulation of virulence genes of the group A streptococcus // J. Bacteriol. - 1997. - 179. - P.5178-5187; Hondorp E.R., Mclver K.S. The Mga virulence regulon: infection where the grass is greener // Mol. Microbiol. - 2007. - 66. - P.1056-1065; Luo F., Lizano S., Banik S., Zhang H., Bessen D.E. Role of Mga in group A streptococcal infection at the skin epithelium // Microb. Pathog. - 2008. - 45. - P.217-224].
Семейство RALP белков-регуляторов транскрипции представлено у S. pyogenes четырьмя белками (RofA, Nra, RALP-3, RALP-4).
К генам факторов патогенности, регулируемым белками семейства RALP, относятся гены MSCRAMMs (гены поверхностных компонентов микробной клетки, распознающих адгезивные молекулы клеточного матрикса), гены белков F1 (prtF1) и F2 (prtF2), связывающих фибронектин, и ген белка, связывающего коллаген (cpa). Под контролем RALP белков также находится экспрессия стрептолизина S (ген sagA), эритрогенного токсина B (ген speB), эритрогенного токсина A (ген speA), глобального регулятора Mga (ген mga) и других [Beckert S. et al. Group A streptococcal rofA gene is involved in the control of several virulence genes and eukaryotic cell attachment and internalization // Infect. Immun. - 2001. - 69. - P.534-537; Podbielski A. et al. Characterization of nra, a global negative regulator gene in group A streptococci // Mol. Microbiol. - 1999. - 31. - P.1051-1064; Kreikemeyer В., Nakata M., Köller Т., Hildisch H., Kourakos V., Standar K., Kawabata S., Glocker M.O., Podbielski A. The Streptococcus pyogenes serotype M49 Nra-Ralp3 transcriptional regulatory network and its control of virulence factor expression from the novel eno ralp3 epf sagA pathogenicity region // Infect. Immun. - 2007. - 75. - P.5698-710].
Белок-регулятор транскрипции Rgg S. pyogenes относится к семейству TIGR01716, согласно базе данных www.jcvi.org. Кроме белка Rgg S. pyogenes к этому семейству относится и ряд других охарактеризованных белков-регуляторов транскрипции грамположительных микроорганизмов, в частности:
- белок Rgg Streptococcus gordonii, регулирующий экспрессию глюкозилтрансферазы GtfG, которая необходима для образования гликанов, играющих важную роль в проявлении микроорганизмом адгезивных свойств и образования бляшки [Sulavik M.C., Tardif G., Clewell D.B. Identification of a gene, rgg, which regulates expression of glucosyltransferase and influences the Spp phenotype of Streptococcus gordonii Challis // J. Bacteriol. - 1992. - 174. - P.3577-3586; Sulavik M.C., Clewell D.B. Rgg is a positive transcriptional regulator of the Streptococcus gordonii gtfG gene // J. Bacteriol. - 1996. - 178. - P.5826-5830];
- белок RovS Streptococcus agalactiae, который регулирует экспрессию супероксиддисмутазы (ген sodA), белка, связывающего фибриноген (ген fbsA), белка-регулятора транскрипции (gbs0230) и белков, кодируемых опероном cyl, участвующих в синтезе гемолизина [Samen U.M., Eikmanns B.J., Reinscheid D.J. The transcriptional regulator RovS controls the attachment of Streptococcus agalactiae to human epithelial cells and the expression of virulence genes // Infect. Immun. - 2006. - 74. - P.5625-5635];
- белок MutR Streptococcus mutans, который регулирует оперон mutAMTFEG, отвечающий за синтез лантибиотика мутацина [Qi F., Chen P., Caufield P.W. Functional analyses of the promoters in the lantibiotic mutacin II biosynthetic locus in Streptococcus mutans // Appl. Environ. Microbiol. - 1999. - 65. - P.652-658];
- белок GadR Lactococcus lactis, активирующий транскрипцию оперона gadBC, который отвечает за глутамат-зависимую толерантность к действию кислот [Sanders J.W., Leenhouts K., Burghoom J., Brands J.R., Venema G., Kok J.. A chloride-inducible acid resistance mechanism in Lactococcus lactis and its regulation // Mol. Microbiol. - 1998. - 27. - P.299-310].
Наиболее изученным из белков-регуляторов семейства TIGR01716 является белок Rgg S. pyogenes. Функциональная активность этого белка была изучена с помощью многочисленных подходов, включая методы молекулярной генетики, биохимии, транскриптомики и протеомики. Так, методом инсерционного мутагенеза ген rgg был инактивирован у штамма S. pyogenes NZ131, и инактивация этого гена привела к изменениям в уровнях транскрипции более 700 генов. Эти изменения отразились на ряде фенотипических свойств S. pyogenes [Chaussee M.S., Ajdic D., Ferretti J.J. The rgg gene of Streptococcus pyogenes positively influences extracellular SPE В production // Infect. Immun. - 1999. - 67. - P.1715-1722; Chaussee M.S. et al. Identification of Rgg-regulated exoproteins of Streptococcus pyogenes // Infect. Immun. - 2001. - 69. - P.822-831; Chaussee M.S., Somerville G.A., Reitzer L. et al. Rgg coordinates virulence factor synthesis and metabolism in Streptococcus pyogenes // J. Bacteriol. - 2003. - 185. - P.6016-6024; Chaussee M.A., Callegari E.A., Chaussee M.S. Rgg regulates growth phase-dependent expression of proteins associated with secondary metabolism and stress in Streptococcus pyogenes // J Bacteriol. - 2004. - 186. - P.7091-7099; Dmitriev A.V. et al. The Rgg regulator of Streptococcus pyogenes influences utilization of nonglucose carbohydrates, prophage induction, and expression of the NAD-glucohydrolase virulence operon // J. Bacteriol. - 2006. - 188. - P.7230-7241], в частности:
1) мутантный штамм оказался неспособен к синтезу эритрогенного токсина B;
2) у мутантного штамма изменилась экспрессия таких факторов патогенности, как стрептолизин O, стрептолизин S, стрептокиназа, белок Маc, НАД-гликогидролаза, M-белок, митогенный фактор MF-1 и др.;
3) инактивация гена rgg оказала влияние на адаптацию S. pyogenes к условиям стресса;
4) инактивация гена rgg повлияла на способность S. pyogenes использовать аминокислоты и углеводы для размножения и роста;
5) инактивация гена rgg повлияла на частоту индукции профага NZ131.1.
Значительные нарушения в экспрессии факторов патогенности (пп.1 и 2) привели к увеличению вирулентных свойств rgg мутантного штамма по сравнению с исходным штаммом NZ131 [Pulliainen A.T., Hytönen J., Haataja S., Finne J. Deficiency of the Rgg regulator promotes H2O2 resistance, AhpCF-mediated H2O2 decomposition, and virulence in Streptococcus pyogenes. J.Bacteriol. - 2008. - 190. - P.3225-3235].
Инактивация гена rgg, исследование регуляторной активности кодируемого им белка Rgg, а также изучение роли белка-регулятора Rgg в проявлении вирулентных свойств S. pyogenes привели к настоящему исследованию. Как приведено выше, инактивации гена rgg вызвала увеличение вирулентных свойств S. pyogenes.
В связи с тем что S. pyogenes является возбудителем большого числа заболеваний, наиболее перспективным является выявление такого гена белка-регулятора, инактивация которого приводила бы не к увеличению, а к снижению или утрате вирулентных свойств.
Задачами данного исследования явились выявление гена-регулятора транскрипции, необходимого для проявления S. pyogenes вирулентных свойств, и инактивация этого гена методом инсерционного мутагенеза с целью получения авирулентного штамма S. pyogenes.
В связи с тем что невозможно предсказать, инактивация какого из генов может привести к снижению/потере или увеличению вирулентных свойств, была разработана единая методологическая схема инактивации генов-регуляторов транскрипции и оценки роли этих генов в метаболизме и вирулентности S. pyogenes.
Для исследования были выбраны несколько штаммов Streptococcus pyogenes: штамм SF370, выделенный от пациента с раневой инфекцией (АТСС 700294, American Type Culture Collection, США), штамм №97, выделенный от пациента с острым тонзиллитом (Россия) и штамм №152 (клинический изолят, Россия).
В качестве первого объекта исследования и, следовательно, первой мишени для инсерционного мутагенеза S. pyogenes был выбран ген mutR, который кодирует белок MutR, относящийся к семейству TIGR01716.
Несмотря на то что S. mutans также имеет ген mutR, последовательность этого гена лишь на 1,4% гомологична последовательности гена mutR S. pyogenes, что свидетельствует о том, что белок MutR S. mutans и белок MutR S. pyogenes являются разными белками.
Следует отметить, что ранее ни ген mutR, ни белок MutR S. pyogenes изучены не были. Ген mutR был лишь охарактеризован как ген предполагаемого активатора транскрипции, имеющий размер 867 п.н. и расположенный на бактериальной хромосоме с 397890 п.н. по 398756 п.н. с нуклеотидной последовательностью, представленной на фиг.1, согласно аннотации к полногеномной последовательности штамма SF370 [Ferretti J.J., McShan W.M., Ajdic D., Savic D.J., Savic G., Lyon K., Primeaux C., Sezate S., Suvorov A.N., Kenton S., Lai H.S, Qian Y., Jia H.G., Najar F.Z., Ren Q., Zhu H., Song L., White J., Yuan X., Clifton S.W., Roe B.A., McLaughlin R. Complete genome sequence of an M1 strain of Streptococcus pyogenes // Proc. Natl. Acad. Sci. USA. - 2001. - 98. - P.4658-4663].
В ходе выполнения исследования уже первая выбранная мишень (ген mutR) оказалась удачной, так как в результате инактивации этого гена был получен авирулентный штамм S. pyogenes SF370, что является доказательством важной роли гена mutR в проявлении микроорганизмом вирулентных свойств. Ген mutR был дополнительно инактивирован еще у двух штаммов S. pyogenes. У одного из этих штаммов инактивация гена mutR также привела к утрате вирулентных свойств - штамм №97[mutR], а у другого - №152[mutR] - вирулентность оказалась значительно снижена по сравнению со штаммом, не подвергшимся мутагенезу с помощью плазмиды pVA891-2[mutR]. Коэффициент летальности для мутантного штамма ниже в 4,7 раза.
Сущностью предлагаемого изобретения является действие на ген-регулятор транскрипции mutR и на проявление Streptococcus pyogenes вирулентных свойств путем направленного мутагенеза (получение авирулентных штаммов этого микроорганизма за счет инактивации гена mutR).
Были получены штаммы S. pyogenes, мутантные по гену mutR, при этом в результате мутации два из трех исследуемых штаммов стали авирулентными, а вирулентные свойства третьего оказались сниженными почти в 5 раз.
Полученные в результате работы штаммы были названы SF370[mutR], №97[mutR] и №152[mutR]. Эти штаммы находятся в коллекции микроорганизмов отдела молекулярной микробиологии Учреждения Российской Академии медицинских наук Научно-исследовательского института экспериментальной медицины Северо-западного отделения РАМН (НИИЭМ СЗО РАМН).
Была выдвинута гипотеза, что полученные авирулентные штаммы могут быть использованы в качестве живой вакцины. Для проверки данного предположения была выполнена серия экспериментов на лабораторных мышах с использованием штамма SF370[mutR]. Мышам вводился авирулентный штамм SF370[mutR], дозы заражения составляли от 1,3×108 КОЕ/животное до 4,4×108 КОЕ/животное, затем через несколько недель (от 2-х до 4-х) им вводился штамм SF370 (доза заражения 3×108 КОЕ/животное). Контрольной группе мышей вместо штамма SF370[mutR] вводился физиологический раствор, затем им так же, как экспериментальной группе, вводился штамм SF370 (доза заражения 3×108 КОЕ/животное).
В результате коэффициент летальности для контрольной группы мышей оказался в 4 раза больше, чем для мышей предварительно вакцинированных авирулентным штаммом. Данный результат подтверждает потенциальную возможность использования авирулентного штамма SF370[mutR] в качестве живой вакцины для профилактики инфекций, вызываемых S. pyogenes.
Получение означенных выше штаммов осуществляли по следующим стадиям.
Инактивация гена регулятора транскрипции mutR Streptococcus pyogenes
Для инактивации гена mutR и получения мутантных штаммов использовали метод инсерционного мутагенеза. Схема эксперимента приведена на фиг.2.
С этой целью PstI-SacI фрагмент гена mutR штамма SF370 клонировали в вектор pVA891-2, содержащий ген устойчивости к эритромицину и неспособный к репликации в грамположительных кокках. Полученная рекомбинантная плазмида размером 3320 п.н. была названа pVA891-2[mutR]. Последующая электропорация штаммов Streptococcus pyogenes SF370, №97 и №152 плазмидой pVA891-2[mutR] приводит к ее встраиванию в ген белка-регулятора mutR и нарушению структурной области гена. Культивирование и селекцию рекомбинантных клонов осуществляли на плотной агаризованной среде или в жидкой среде на основе Todd-Hewitt Broth (HiMedia, Индия), содержащей эритромицин (2,5 мкг/мл). В результате электропорации штаммов Streptococcus pyogenes SF370, №97 и №152 были получены клоны, устойчивые к действию эритромицина.
Анализ полученных клонов на наличие плазмиды pVA891-2[mutR] в гене mutR
Наличие плазмиды pVA891-2 [mutR], встроенной в структурную область гена mutR, выявляли методами полимеразной цепной реакции (ПЦР) и секвенирования.
1. Анализ методом полимеразной цепной реакции
Наличие плазмиды pVA891-2[mutR] в гене mutR определяли методом ПЦР у рекомбинантных клонов, сконструированных на основе всех трех изученных штаммов (SF370, №97 и №152). Подробное описание приводится для анализа рекомбинантных клонов штамма SF370, а рекомбинантные клоны штаммов №97 и №152 исследовали аналогичным образом.
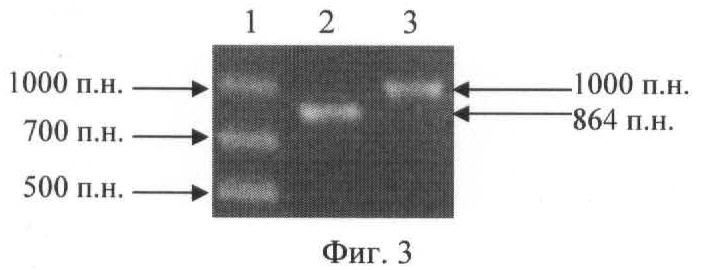
Наличие плазмиды pVA891-2[mutR] в гене mutR определяли при использовании двух пар праймеров: MutR-1; 40/1 и MutR+1; 40/2. Как видно из фиг.2, каждая из этих пар праймеров может привести к амплификации фрагмента ДНК только на матрице хромосомной ДНК мутантного штамма. Ожидаемый размер фрагментов составляет:
- 864 п.н. (при использовании праймеров MutR-1 и 40/1);
- 1000 п.н. (при использовании праймеров MutR+1 и 40/2).
Результаты ПЦР приведены на фиг.3. Анализ клона, устойчивого к действию эритромицина, методом ПЦР, где трек 1 - маркер молекулярных весов; трек 2 - ПЦР с праймерами MutR-1, 40/1; трек 3 - ПЦР с праймерами MutR+1, 40/2. Как видно из фиг.3, в результате анализа исследуемого клона были получены амплификаты ожидаемых размеров при использовании обеих пар праймеров. В качестве отрицательного контроля был использован штамм SF370, на матрице ДНК которого ПЦР с парами праймеров MutR-1; 40/1 и MutR+1; 40/2 не должна была приводить к образованию продуктов амплификации (фиг.2). Как и ожидалось, ПЦР реакция с этими парами праймеров не привела к получению продуктов амплификации.
Кроме того, на матрицах ДНК штамма SF370 и клона, устойчивого к действию эритромицина, при использовании Phusion Flash High-Fidelity PCR Master Mix (Finnzymes, Finland) была проведена ПЦР с праймерами MutR-1, MutR+1. Для штамма SF370 размер полученного амплификата составил 1126 п.н., а для рекомбинантого клона ~4500 п.н., что доказало наличие в гене mutR вставки размером 3320 п.н., что соответствует размеру плазмиды pVA891-2[mutR] (фиг.4 - ПЦР-анализ штамма SF370 (А) и рекомбинантного клона (Б) с праймерами MutR-1, MutR+1 (трек 2), трек 1 - маркер молекулярных весов).
Аналогичные результаты были получены и при анализе штаммов №97, №152 и сконструированных на их основе рекомбинантных клонов.
Таким образом, полученные результаты позволяют сделать вывод, что рекомбинантные клоны, сконструированные на основе штаммов SF370, №97 и №152 являются мутантными штаммами SF370[mutR], №97[mutR] и №152[mutR] соответственно, а встраивание плазмиды pVA891-2 [mutR] произошло, как и ожидалось, в структурную область гена mutR.
2. Анализ методом секвенирования
Результаты метода ПЦР являются достаточными для подтверждения искомой мутации в гене mutR штаммов SF370, №97 и №152. Тем не менее, для того чтобы дополнительно подтвердить, что встраивание плазмиды pVA891-2[mutR] привело к получению ожидаемой генетической конструкции в области гена mutR, амплификаты, полученные на матрице ДНК одного из штаммов (SF370[mutR]) с использованием пар праймеров MutR-1; 40/1 и MutR+1; 40/2, были секвенированы. Результаты секвенирования приведены на фиг.5 Нуклеотидные последовательности амплификатов, полученных с парами праймеров MutR-1; 40/1 (А) и MutR+1; 40/2 (Б), жирным шрифтом выделены фрагменты гена mutR; старт-кодон ATG и стоп-кодон TGA гена mutR соответствуют нуклеотидам 397890-397892 и 398754-398756, согласно полногеномной последовательности штамма SF370 [Ferretti J.J., McShan W.M., Ajdic D., Savic D.J., Savic G., Lyon K., Primeaux C., Sezate S., Suvorov A.N., Kenton S., Lai H.S., Lin S.P., Qian Y., Jia H.G., Najar F.Z., Ren Q., Zhu H., Song L., White J., Yuan X., Clifton S.W., Roe B.A., McLaughlin R. Complete genome sequence of an Ml strain of Streptococcus pyogenes // Proc. Natl. Acad. Sci. USA. - 2001. - 98. - P.4658-4663]. Таким образом, нуклеотидная последовательность и структура генетической конструкции в области гена mutR в мутантном штамме полностью соответствует ожидаемой, приведенной на фиг.2.
Влияние инактивации гена mutR на свойства S. pyogenes
Сравнительный анализ штаммов SF370 и SF370[mutR] проводили по следующим признакам:
- основные морфологические свойства микроорганизма;
- динамика роста в обогащенной среде Todd-Hewitt Broth;
- белковый состав клеточных лизатов и секретируемых белков.
Такие морфологические свойства, как размер колоний, диаметр зоны гемолиза и характер роста на плотной кровяной агаризованной среде, были сходными у штаммов SF370 и SF370[mutR]. Результаты микроскопии штаммов дополнительно показали, что инактивация гена mutR не привела к изменениям в диаметре клеток и количестве клеток в цепочках.
При росте штаммов в жидкой питательной среде Todd-Hewitt Broth были отмечены существенные различия. Штамм SF370 имел характерный придонный рост, а штамм SF370[mutR] рос во всем объеме среды, и только к середине стационарной фазы его рост становился придонным и сходным с таковым штамма SF370. При этом инактивация гена mutR привела к значительным изменениям в скорости роста штамма и продолжительности лаг- и лог-фаз роста. Максимальные значения OD600 для штаммов SF370[mutR] и SF370 значительно отличались и составили 1,1 и 0,8 соответственно.
Анализ штамма S. pyogenes SF370 и штамма SF370[mutR] с помощью SDS-PAGE показал различия в составе/количестве секретируемых белков. В частности, только у штамма SF370[mutR} присутствовал фрагмент размером 74 кДа. Кроме того, штаммы отличались и количеством белков, составляющих белковые полосы размером 34 кДа. Анализ белковых полос размерами 34 кДа у обоих штаммов методом масс-спектрометрии показал, что каждая из этих полос представлена только белком Sic, а его экспрессия существенно выше у штамма SF370[mutR] по сравнению со штаммом SF370.
Штаммы №97 и №152, а также их мутанты по гену mutR были исследованы на уровни экспрессии секретируемых нуклеаз. Было показано, что экспрессия секретируемых нуклеаз у мутантных штаммов №97[mutR] и №152[mutR] существенно ниже по сравнению со штаммами №97 и №152. При сравнение уровня экспрессии секретируемых нуклеаз у штаммов SF370 и SF370[mutR] существенной разницы выявлено не было. Фиг.6 - экспрессия секретируемых нуклеаз.
Оценка вирулентных свойств
Для оценки вирулентных свойств изучаемых штаммов применяли внутрибрюшинное заражение лабораторных мышей и метод прямого бактерицидного теста.
1. Внутрибрюшинное заражение
В качестве лабораторных животных использовали беспородных мышей (самцы, 10-12 нед, вес 14-16 г, питомник лабораторных животных РАМН «Рапполово»). Для моделирования стрептококковой инфекции штаммы выращивали при 37°C в 40 мл бульона Todd-Hewitt Broth в течение ночи, клетки центрифугировали и отмывали физиологическим раствором. Отмытые клетки ресуспендировали в физиологическом растворе. Животным вводили внутрибрюшинно по 0,5 мл микробной суспензии с различными концентрациями возбудителя, в зависимости от штамма микроорганизма (дозы заражения 1×108 и 5×108 КОЕ/животное для штаммов SF370 и SF370[mutR], 3,5×108 КОЕ/животное для штаммов №97 и №97[mutR] и 4×108 КОЕ/животное для штаммов №152 и №152[mutR]). Для каждой дозы заражения использовали по 15 мышей. Наблюдение вели в течение 10 дней. Для подтверждения гибели животных от стрептококковой инфекции делали счетные высевы микроорганизмов из селезенок.
Вирулентность штаммов оценивали по коэффициенту летальности - KL.
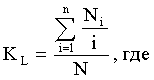
Ni - количество мышей, павших в i-ые сутки;
N - общее число животных, использованных для анализа данной дозы заражения.
Для штамма SF370 при дозах заражения 1×108 и 5×108 значение коэффициента летальности составило 0,13 и 0,54 соответственно. Мутантный штамм SF370[mutR] при этих же дозах оказался авирулентным, не приведя к гибели животных, фиг.7 Динамика гибели лабораторных мышей при внутрибрюшинном заражении штаммами SF370 и SF370[mutR] (А), штаммами №97 и №97[mutR] (Б) и штаммами №152 и №52[mutR] (В).
Для штамма №97 при дозе заражения 3,5×108 значение коэффициента летальности составило 0,26. Мутантный штамм №97[mutR] оказался авирулентным, не приведя к гибели животных (фиг.7Б).
Для штаммов №152 и №152[mutR] при дозе заражения 4×108 КОЕ/животное значение коэффициентов летальности составило 0,56 и 0,15 соответственно, свидетельствуя о значительном снижении вирулентных свойств в результате инактивации гена mutR (фиг.7В).
2. Прямой бактерицидный тест
Для проведения прямого бактерицидного теста 100 мкл физиологического раствора, содержащих 3×102 КОЕ S. pyogenes, смешивали с 300 мкл цельной человеческой крови и инкубировали при 37°C, встряхивая каждые 20 минут. Пробы отбирали каждый час в течение 6 часов. Количество жизнеспособных стрептококковых КОЕ определяли с помощью счетных высевов.
Методом прямого бактерицидного теста были проанализированы штаммы SF370 и SF370[mutR].
В результате исследований было обнаружено, что в цельной человеческой крови исходный штамм SF370 интенсивно размножался, а штамм SF370[mutR] погибал (фиг.8. Результаты прямого бактерицидного теста).
Таким образом, обоими методами (моделирование стрептококковой инфекции на лабораторных животных и метод прямого бактерицидного теста в человеческой крови) показано, что инактивация гена mutR приводит либо к полной потере вирулентных свойств у штаммов S. pyogenes SF370[mutR] и №97[mutR], либо к сильному снижению вирулентных свойств у штамма №152[mutR].
В результате инактивации гена mutR S. pyogenes было получено два авирулентных штамма, SF370[mutR] и №97[mutR], для штамма SF370[mutR] показана возможность использования его в качестве живой вакцины.


{kind=link}